2742064
Beschreibung
Mindmap von Catherine Salmon, aktualisiert more than 1 year ago
|
|
Erstellt von Catherine Salmon
vor mehr als 9 Jahre
|
|
Mechanisms of Synaptic Plasticity
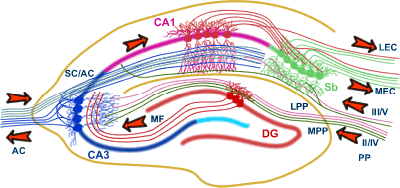
- Hippocampus
- located within the medial temporal lobe of the brain. It
forms part of the limbic system
- Hippocampal Pathways
- Hippocampal Function
- CA1 region of the hippocampus is vital for some forms of learning
- Hippocampus is responsible for the acquisition of
memory with particular emphasis on spatial memory
- maintains a neural representation of spatial environment
- maintains a neural representation of spatial environment
- CA1 region of the hippocampus is vital for some forms of learning
- located within the medial temporal lobe of the brain. It
forms part of the limbic system
- Presynaptic mechanisms of plasticity
- Short-term Plasticity
- 3 types of Short-term plasticity
- Paired Pulse Depression
- Synapses that release transmitter at a high
release probability
- 3 factors for reduced release
- Depletion of the proportion of vesicles that
are in the Readily Releasable Pool (RRP)
- Release site innactivation due to vesicular membrane proteins
interfering with membrane consistency/ protein interactions
- Decreased calcium influx. Decreased activity of Ca/
Calmodulin complex. P-type Ca Channels not activated
- Depletion of the proportion of vesicles that
are in the Readily Releasable Pool (RRP)
- Synapses that release transmitter at a high
release probability
- Paired Pulse Potentiation
- Paired Pulse Facilitation
- Hippocampal mossy fibres participate in a lot
of PPF at the synapse to CA3 Pyramidal cells.
- Low Release probability
- Metabotropic receptors are the "brakes" the cell applies to ensure that
there is a limit to the magnitude of frequency-dependent facillitation
- Fibres contain adenosine A1 GPCRs on the presynaptic cell. These
receptors have an inhibitory effect on glutamate release
- Mossy fibres also express metabatropic Glutamate receptors (mGluR 2 or 3) which
also suppress glutamate release at the synapse by inhibiting calcium entry
- negative feedback to stop over excitation/ glutamate
release as Glutamate spills over to activate mGluR
- Activation of G protein-coupled receptors can transiently inhibit vesicular release through the
release of Gβγ which binds to both voltage-dependent calcium channels to reduce calcium influx, and
directly to the C-terminus region of the SNARE protein SNAP-25
- negative feedback to stop over excitation/ glutamate
release as Glutamate spills over to activate mGluR
- Fibres contain adenosine A1 GPCRs on the presynaptic cell. These
receptors have an inhibitory effect on glutamate release
- Low Release probability
- Hippocampal mossy fibres participate in a lot
of PPF at the synapse to CA3 Pyramidal cells.
- Pairs of pulses delivered at
intervals of several tens of
milliseconds
- Paired Pulse Depression
- Unlike LTD or LTP, the effects last for a
maximum of a couple of seconds
- Calcium dependency
- Short-term facilitation
- Transmitter release
- Transient increases in [Ca2+]i, in close proximity to
voltage-gated Ca channels, drive phasic transmitter
release. This contributes to depolarisation
- Ca2+ channels will remain open until repolarisation occurs
- Calcium signal will reduce quickly due to diffusion
- Residual Calcium signals remain (at a concentration of ~500nM) as Calcuim
buffering and extrusion (active transport out of the cell or into intracellular
stores) are slow therefore there is a slow signal decay
- Residual calcium builds up during repetitive stimulation (5-10Hz)
- Asynchronous signalling
- Synchrony
- Synchrony
- Residual calcium may serve as a mechanism for
short-term plasticity within neuronal circuits
- Asynchronous signalling
- Residual calcium builds up during repetitive stimulation (5-10Hz)
- Residual Calcium signals remain (at a concentration of ~500nM) as Calcuim
buffering and extrusion (active transport out of the cell or into intracellular
stores) are slow therefore there is a slow signal decay
- Calcium signal will reduce quickly due to diffusion
- [Ca2+] levels in the tens of micromolar range
- Ca2+ channels will remain open until repolarisation occurs
- Mechanism of transmitter release
- How many molecules of Ca2+ are needed for Synaptotagmin to perform vesicular exocytosis??
- Ca sensitive protein which, when bound to embedded
proteins, forms the SNARE complex
- 2 models
- 5-site
- Does not account for the calcuim concentration range at
which exocytosis of vesicles occurs
- may therefore require activation at an allosteric site in low calcium concentrations
- may therefore require activation at an allosteric site in low calcium concentrations
- Does not account for the calcuim concentration range at
which exocytosis of vesicles occurs
- 2-site 5-site
- 2-site filled
- Possible at low concentrations of calcium, slow calcium sensor
- Asynchronous
- Possible at low concentrations of calcium, slow calcium sensor
- 5-site filled
- only possible at high concentrations, fast calcium sensor
- Synchronus
- only possible at high concentrations, fast calcium sensor
- 2-site filled
- 5-site
- Ca sensitive protein which, when bound to embedded
proteins, forms the SNARE complex
- How many molecules of Ca2+ are needed for Synaptotagmin to perform vesicular exocytosis??
- Transient increases in [Ca2+]i, in close proximity to
voltage-gated Ca channels, drive phasic transmitter
release. This contributes to depolarisation
- Synchrony
- Short-term facilitation
- 3 types of Short-term plasticity
- Long-term
- mGluR may also have a long-term presynaptic effect
- Activation of G protein-coupled receptors can transiently inhibit vesicular release through the
release of Gβγ which binds to both voltage-dependent calcium channels to reduce calcium influx, and
directly to the C-terminus region of the SNARE protein SNAP-25.
- Alford et al. 2013 showed that it is necessary but not sufficient to produce LTD alone
- Alford et al. 2013 showed that it is necessary but not sufficient to produce LTD alone
- Activation of G protein-coupled receptors can transiently inhibit vesicular release through the
release of Gβγ which binds to both voltage-dependent calcium channels to reduce calcium influx, and
directly to the C-terminus region of the SNARE protein SNAP-25.
- Presynaptic LTP in the Mossy fibre/ CA3 pyramidal synapse
- Increases in PRESYNAPTIC Calcium
- post-synaptic calcium has a very small role
- N-type, P/Q-type and R-type Calcium Channels have all been
implemented as important for calcium entry
- N or P/Q type are sufficient for LTP induction
- Access to AC and release machinery
- Access to AC and release machinery
- R-type channels
- Lower threshold for LTP induction
- Does not have access to the release machinery
- Preferential access to
Adenylyl Cyclase (AC)
- Through
Calmodulin/Ca
complex
- Through
Calmodulin/Ca
complex
- Lower threshold for LTP induction
- Amplitude of EPSP does not change significantly between
WT and mice with a R-type subunit knockout
- Enhancement of neurotransmitter release involves enhanced Ca coupling to SNARE complex
- Enhancement of neurotransmitter release involves enhanced Ca coupling to SNARE complex
- N or P/Q type are sufficient for LTP induction
- post-synaptic calcium has a very small role
- Increases in PRESYNAPTIC Calcium
- mGluR may also have a long-term presynaptic effect
- Short-term Plasticity
- Post synaptic Mechanisms of Plasticity
- Long Term Potentiation (LTP)
- Molecular mechanisms
of long-term
potentiation
- Signalling Cascades
- Increased [Ca2+]i allows for Calmodulin to bind 4Ca2+ cations
- Ca/Calmodulin complex activates CAMKII (Cacium/calmodulin kinase 2) ***
- Ca/Calmodulin complex also activates membrane-bound Adenylyl Cyclase
- ATP > cAMP
- cAMP bind and activates Phosphokinase A (PKA)
- PKA traslocates into the nucleus
- Kinases Activate CREB1 (cAMP response element binding protein)
- CREB activates CRE (cAMP response element)
- Calcium-regulated gene expression of certain growth factors that may
enable De novo spine formation
- Calcium-regulated gene expression of certain growth factors that may
enable De novo spine formation
- CREB activates CRE (cAMP response element)
- Kinases Activate CREB1 (cAMP response element binding protein)
- PKA can also activate MAPK (mitogen activated protein kinase)
- MAPK translocates into the nucleus
- MAPK translocates into the nucleus
- PKA traslocates into the nucleus
- cAMP bind and activates Phosphokinase A (PKA)
- ATP > cAMP
- Ca/Calmodulin complex activates CAMKII (Cacium/calmodulin kinase 2) ***
- Increased [Ca2+]i allows for Calmodulin to bind 4Ca2+ cations
- Signal transduction
- NMDA receptors
- NMDA receptors open with Glutamate
binding on the extracellular domain.
- "coincidence detectors" as require Mg
block removal and glutamate
- "coincidence detectors" as require Mg
block removal and glutamate
- Influx of Ca2+, Na+ and efflux of K+
- Intracellular Ca2+ increases with opening of
NMDA channels
- Intracellular Ca2+ increases with opening of
NMDA channels
- NMDA receptors open with Glutamate
binding on the extracellular domain.
- AMPA Receptors
- On ligand (aglutamate) binding,
AMPA receptors open, allowing for
sodium influx and potassium eflux
- Intracellular concentrations of Na are higher than
external concentrations. The net possitive charge
intracellularly resultis in the magnesium block in
NMDA channels to be removed
- Increased [Ca2+]i allows for the Ca/Calmodulin complex to form
- Ca/Calmodulin activates CAMKII by phosphorylation ***
- CAMKII phosphorylates Stargazin scaffold protein
- PSD95 can therefore bind and can relocate the AMPA receptor to the
post-synaptic density
- PSD95 can therefore bind and can relocate the AMPA receptor to the
post-synaptic density
- CAMKII also phosphorylates the GluR1
subunit at Serine 831
- Increases the Conductance of the AMPA Receptor
- Increases the Conductance of the AMPA Receptor
- CAMKII phosphorylates Stargazin scaffold protein
- Ca/Calmodulin activates CAMKII by phosphorylation ***
- Intracellular concentrations of Na are higher than
external concentrations. The net possitive charge
intracellularly resultis in the magnesium block in
NMDA channels to be removed
- On ligand (aglutamate) binding,
AMPA receptors open, allowing for
sodium influx and potassium eflux
- NMDA receptors
- Signalling Cascades
- How is LTP induced?
- Pyramidal neurons fire at a frequency of 5Hz. This is known as "theta rhythm"
- Theta rhythm
- Three trains of stimuli that are 20 seconds apart
- Each train composed of 10 stimulus epochs which are delivered at 5Hz (i.e. 200ms apart)
- each epoch is made up of four individual stimuli at 100Hz
- each epoch is made up of four individual stimuli at 100Hz
- Each train composed of 10 stimulus epochs which are delivered at 5Hz (i.e. 200ms apart)
- Three trains of stimuli that are 20 seconds apart
- during exploration (spatial mapping)
- Theta rhythm
- Pyramidal neurons fire at a frequency of 5Hz. This is known as "theta rhythm"
- Discovery
- Raising the frequency of firin from 0.5Hz to 15Hz
- follow grey lines
- Baseline at 0.5 Hz
- Initial facilitation
of EPSP at 15 Hz
- Rapid decline past
baseline
- Gradual rise to an
increased baseline mV
- Gradual rise to an
increased baseline mV
- Rapid decline past
baseline
- Initial facilitation
of EPSP at 15 Hz
- Baseline at 0.5 Hz
- follow grey lines
- Raising the frequency of firin from 0.5Hz to 15Hz
- Properties
- 4 major properties/ conditions neccessary for LTP
- Cooperativity
- LTP can be induced by a single, strong, tetanic signal to a synapse
OR via weaker stimulation of may cooperating pathways
- LTP can be induced by a single, strong, tetanic signal to a synapse
OR via weaker stimulation of may cooperating pathways
- Associativity
- the contributing fibres need to be active together
- information from the two contributing sources must coincide
both temporally and spatially
- Weak tetanic stimulus in one location will be
insufficient in producing LTP, but paired with a
strong signal in another location can induce LTP
- information from the two contributing sources must coincide
both temporally and spatially
- the contributing fibres need to be active together
- Input specificty
- LTP will only occur at synapses which receive tetanic stimulation,
even if it is within the same cell, other synapses which do not
recieve stimulation will not share in synaptic enhancement (i.e.
increased expression of AMPA or NMDA receptors)
- LTP will only occur at synapses which receive tetanic stimulation,
even if it is within the same cell, other synapses which do not
recieve stimulation will not share in synaptic enhancement (i.e.
increased expression of AMPA or NMDA receptors)
- Persistance
- LTP is also persistent, it can last for several hours and even for many
months suggesting that a sustained physiological change
accompanies memory formation
- LTP is also persistent, it can last for several hours and even for many
months suggesting that a sustained physiological change
accompanies memory formation
- Cooperativity
- 4 major properties/ conditions neccessary for LTP
- 'Hebb Rule' = if both pre and post-synaptic cell are excited at the same time, the synaptic connection is strengthened
- Molecular mechanisms
of long-term
potentiation
- Glutamate is the main excitatory
neurotransmitter in the synaptic transmission
between Schaffer Collateral fibres and the CA1
pyramidal cells
- Long Term Potentiation (LTP)
Medienanhänge
{kind=link}
{kind=link}
Möchten Sie kostenlos Ihre eigenen Mindmaps mit GoConqr erstellen? Mehr erfahren.