3613691

Ciclo de Krebs
- ciclo metabólico de importancia fundamental en
todas las células que utilizan oxígeno durante el
proceso de respiración celular
- En estos organismos aeróbicos, el ciclo de Krebs
es el anillo de conjunción de las rutas metabólicas
responsables de la degradación y desasimilación
de los carbohidratos, las grasas y las proteínas en
anhídrido carbónico y agua, con la formación de
energía química.
- En estos organismos aeróbicos, el ciclo de Krebs
es el anillo de conjunción de las rutas metabólicas
responsables de la degradación y desasimilación
de los carbohidratos, las grasas y las proteínas en
anhídrido carbónico y agua, con la formación de
energía química.
- Etapas
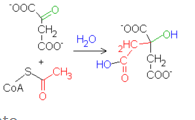
- Reacción 1: Citrato sintasa (De oxalacetato a citrato
- El sitio activo de la enzima, activa el acetil-CoA para
hacerlo afín a un centro carbonoso del oxalacetato.
- el grupo tioéster (CoA) se hidroliza, formando así la
molécula de citrato.
- La reacción es sumamente exoergónica (ΔG'°=-31.4 kJ/mol),
motivo por el cual este paso es irreversible.
- El citrato producido por la enzima, además, es capaz de
inhibir competitivamente la actividad de la enzima.
- permite una completa regulación del ciclo de Krebs completo, convirtiendo a la
enzima en una especie de marcapasos del ciclo
- permite una completa regulación del ciclo de Krebs completo, convirtiendo a la
enzima en una especie de marcapasos del ciclo
- El citrato producido por la enzima, además, es capaz de
inhibir competitivamente la actividad de la enzima.
- La reacción es sumamente exoergónica (ΔG'°=-31.4 kJ/mol),
motivo por el cual este paso es irreversible.
- el grupo tioéster (CoA) se hidroliza, formando así la
molécula de citrato.
- El sitio activo de la enzima, activa el acetil-CoA para
hacerlo afín a un centro carbonoso del oxalacetato.
- Reacción 2: Aconitasa (De citrato a isocitrato)
- La aconitasa cataliza la isomerización del citrato a isocitrato, por
la formación de cis-aconitato
- La enzima cataliza también la reacción inversa,
pero en el ciclo de Krebs tal reacción es
unidireccional a causa de la ley de acción de masa
- las concentraciones (en condiciones
estándar) de citrato (91%), del
intermediario cis-aconitato (3%) y de
isocitrato (6%), empujan decididamente
la reacción hacia la producción de
isocitrato
- En el sitio activo de la enzima está presente un clúster hierro-azufre
que, junto a algunos residuos de aminoácidos polares, liga el sustrato
- la unión al sustrato se asegura por la presencia de un resto de
serina, de arginina, de histidina y de aspartato, que permiten sólo
la unión estereospecifica del citrato 1R,2S, rechazando la forma
opuesta.
- la unión al sustrato se asegura por la presencia de un resto de
serina, de arginina, de histidina y de aspartato, que permiten sólo
la unión estereospecifica del citrato 1R,2S, rechazando la forma
opuesta.
- las concentraciones (en condiciones
estándar) de citrato (91%), del
intermediario cis-aconitato (3%) y de
isocitrato (6%), empujan decididamente
la reacción hacia la producción de
isocitrato
- La enzima cataliza también la reacción inversa,
pero en el ciclo de Krebs tal reacción es
unidireccional a causa de la ley de acción de masa
- La aconitasa cataliza la isomerización del citrato a isocitrato, por
la formación de cis-aconitato
- Reacción 3: Isocitrato deshidrogenasa (De isocitrato
a oxoglutarato)
- La isocitrato deshidrogenasa mitocondrial es una
enzima dependiente de la presencia de NAD+ y de
Mn2+ o Mg2+
- la presencia de un ión bivalente, que forma un
complejo con los oxígenos del grupo carboxilo
en posición alfa, aumenta la electronegatividad
de esa región molecular
- salida de una molécula de CO2, que conduce a la
formación de α-cetoglutarato, caracterizado por dos
carboxilos en las extremidades y una cetona en posición
alfa con respecto de uno de los dos grupos carboxilo.
- salida de una molécula de CO2, que conduce a la
formación de α-cetoglutarato, caracterizado por dos
carboxilos en las extremidades y una cetona en posición
alfa con respecto de uno de los dos grupos carboxilo.
- la presencia de un ión bivalente, que forma un
complejo con los oxígenos del grupo carboxilo
en posición alfa, aumenta la electronegatividad
de esa región molecular
- La isocitrato deshidrogenasa mitocondrial es una
enzima dependiente de la presencia de NAD+ y de
Mn2+ o Mg2+
- Reacción 4: α-cetoglutarato
deshidrogenasa (De
oxoglutarato a Succinil-CoA)
- Después de la conversión del isocitrato en
α-cetoglutarato se produce una segunda
reacción de descarboxilación oxidativa, que
lleva a la formación de succinil CoA
- alfa-cetoglutarato deshidrogenasa está compuesta
de tres enzimas diferentes:
- Subunidad E1: las dos cetoglutarato deshidrogenasas.
Subunidad E2: la transuccinilasa. (La subunidad E1 y E2
presentan una gran homología con las de la piruvato
deshidrogenasa.) * Subunidad E3: la dihidrolipoamida
deshidrogenasa, que es el mismo polipéptido presente
en el otro complejo enzimático.
- Subunidad E1: las dos cetoglutarato deshidrogenasas.
Subunidad E2: la transuccinilasa. (La subunidad E1 y E2
presentan una gran homología con las de la piruvato
deshidrogenasa.) * Subunidad E3: la dihidrolipoamida
deshidrogenasa, que es el mismo polipéptido presente
en el otro complejo enzimático.
- alfa-cetoglutarato deshidrogenasa está compuesta
de tres enzimas diferentes:
- Después de la conversión del isocitrato en
α-cetoglutarato se produce una segunda
reacción de descarboxilación oxidativa, que
lleva a la formación de succinil CoA
- Reacción 5: Succinil-CoA sintetasa
(De Succinil-CoA a succinato)
- La citrato sintasa se sirve de un
intermediario con tal unión a alta
energía para llevar a cabo la fusión entre
una molécula con dos átomos de
carbono (acetil-CoA
- La enzima succinil-CoA sintetasa se sirve de tal
energía para fosforilar un nucleósido difosfato
purinico como el GDP.
- El primer paso de la reacción genera un nuevo
intermediario a alta energía, conocido como succinil
fosfato.
- El primer paso de la reacción genera un nuevo
intermediario a alta energía, conocido como succinil
fosfato.
- La enzima succinil-CoA sintetasa se sirve de tal
energía para fosforilar un nucleósido difosfato
purinico como el GDP.
- Una histidina presente en el sitio catalítico
remueve el fosfato de la molécula glucídica,
generando el producto succinato y una
molécula de fosfohistidina, que dona
velozmente el fosfato a un nucleósido
difosfato, recargándolo a trifosfato
- sencialmente trasladar grupos fosfato hacia el
ATP, en una reacción catalizada por la enzima
nucleósido difosfoquinasa.
- sencialmente trasladar grupos fosfato hacia el
ATP, en una reacción catalizada por la enzima
nucleósido difosfoquinasa.
- La citrato sintasa se sirve de un
intermediario con tal unión a alta
energía para llevar a cabo la fusión entre
una molécula con dos átomos de
carbono (acetil-CoA
- Reacción 6:
Succinato
deshidrogenasa
(De succinato a
fumarato
- La parte final del ciclo consiste en la reorganización de
moléculas a cuatro átomos de carbono hasta la
regeneración del oxalacetato
- Para que eso sea posible, el grupo metilo presente
en el succinato tiene que convertirse en un
carbonilo
- extracción ulterior de energía mediante
la formación de FADH2 y NADH.
- La primera reacción de oxidación es catalizada por el
complejo enzimático de la succinato deshidrogenasa
- única enzima del ciclo que tiene como aceptor de hidrógeno al FAD en
vez de al NAD+. El FAD es enlazado de modo covalente a la enzima por un
residuo de histidina
- única enzima del ciclo que tiene como aceptor de
hidrógeno al FAD en vez de al NAD+. El FAD es
enlazado de modo covalente a la enzima por un
residuo de histidina
- Tal posición se debe a la implicación de la enzima en la cadena
de transporte de los electrones.
- Los electrones pasados sobre el FAD se introducen
directamente en la cadena gracias a la unión estable entre la
enzima y el cofactor mismo
- Los electrones pasados sobre el FAD se introducen
directamente en la cadena gracias a la unión estable entre la
enzima y el cofactor mismo
- Tal posición se debe a la implicación de la enzima en la cadena
de transporte de los electrones.
- única enzima del ciclo que tiene como aceptor de
hidrógeno al FAD en vez de al NAD+. El FAD es
enlazado de modo covalente a la enzima por un
residuo de histidina
- única enzima del ciclo que tiene como aceptor de hidrógeno al FAD en
vez de al NAD+. El FAD es enlazado de modo covalente a la enzima por un
residuo de histidina
- La primera reacción de oxidación es catalizada por el
complejo enzimático de la succinato deshidrogenasa
- extracción ulterior de energía mediante
la formación de FADH2 y NADH.
- Para que eso sea posible, el grupo metilo presente
en el succinato tiene que convertirse en un
carbonilo
- La parte final del ciclo consiste en la reorganización de
moléculas a cuatro átomos de carbono hasta la
regeneración del oxalacetato
- Reacción 7: Fumarasa (De fumarato a L-malato)
- La fumarasa cataliza la adición en trans de un protón y un
grupo OH- procedentes de una molécula de agua
- La hidratación del fumarato produce L-malato.
- La hidratación del fumarato produce L-malato.
- La fumarasa cataliza la adición en trans de un protón y un
grupo OH- procedentes de una molécula de agua
- Reacción 8: Malato deshidrogenasa (De
L-malato a oxalacetato)
- La última reacción del ciclo de Krebs
consiste en la oxidación del malato a
oxalacetat
- La reacción, catalizada por la malato
deshidrogenasa, utiliza otra molécula
de NAD+ como aceptor de hidrógeno,
produciendo NADH.
- La energía libre de Gibbs asociada
con esta última reacción es
decididamente positiva, a
diferencia de las otras del ciclo
- La actividad de la enzima es remolcada por el consumo de oxalacetato por
parte de la citrato sintasa, y de NADH por parte de la cadena de transporte de
electrones.
- La actividad de la enzima es remolcada por el consumo de oxalacetato por
parte de la citrato sintasa, y de NADH por parte de la cadena de transporte de
electrones.
- La energía libre de Gibbs asociada
con esta última reacción es
decididamente positiva, a
diferencia de las otras del ciclo
- La reacción, catalizada por la malato
deshidrogenasa, utiliza otra molécula
de NAD+ como aceptor de hidrógeno,
produciendo NADH.
- La última reacción del ciclo de Krebs
consiste en la oxidación del malato a
oxalacetat
- Reacción 1: Citrato sintasa (De oxalacetato a citrato
- otras rutas metabólicas
- Catabolismo de los carbohidratos
- La glucolisis degrada la glucosa ) en piruvato y un α-cetoácido
que contiene tres átomos de carbono
- La glucolisis degrada la glucosa (y otras moléculas de seis átomos de
carbono) en piruvato y un α-cetoácido que contiene tres átomos de
carbono
- pierde un átomo de carbono y se convierte en acetil-CoA mediante la piruvato desihdrogenasa
- En el interior de la mitocondria, el acetil-CoA puede entrar en el ciclo de Krebs, como se describió
anteriormente
- En el interior de la mitocondria, el acetil-CoA puede entrar en el ciclo de Krebs, como se describió
anteriormente
- pierde un átomo de carbono y se convierte en acetil-CoA mediante la piruvato desihdrogenasa
- La glucolisis degrada la glucosa (y otras moléculas de seis átomos de
carbono) en piruvato y un α-cetoácido que contiene tres átomos de
carbono
- La glucolisis degrada la glucosa ) en piruvato y un α-cetoácido
que contiene tres átomos de carbono
- Catabolismo de las proteínas
- En lo que concierne a las proteínas,
son degradadas mediante
mecanismos de proteolisis por
enzimas proteasas, que las trocean en
sus constituyentes fundamentales: los
aminoácidos
- Algunos aminoácidos pueden constituir una fuente de energía, ya que
son convertibles en intermediarios del ciclo mismo, por ejemplo el
aspartato, la valina y la isoleucina
- Otros, convertibles en moléculas glucídicas, pueden entrar en el ciclo pasando
por las rutas catabólicas típicas de los glúcidos, por ejemplo la alanina,
convertible en piruvato.
- Otros, convertibles en moléculas glucídicas, pueden entrar en el ciclo pasando
por las rutas catabólicas típicas de los glúcidos, por ejemplo la alanina,
convertible en piruvato.
- Algunos aminoácidos pueden constituir una fuente de energía, ya que
son convertibles en intermediarios del ciclo mismo, por ejemplo el
aspartato, la valina y la isoleucina
- En lo que concierne a las proteínas,
son degradadas mediante
mecanismos de proteolisis por
enzimas proteasas, que las trocean en
sus constituyentes fundamentales: los
aminoácidos
- Catabolismo de los lípidos
- En el catabolismo de los lípidos, los triglicéridos
son hidrolizados por enzimas lipasas para formar
ácidos grasos y glicero
- En los organismos superiores,
el glicerol puede entrar en la
glucolisis a nivel hepático o ser
transformado en glucosa a
través de la hidroxiacetona
fosfato y el
gliceraldehído-3-fosfato,
siguiendo la ruta metabólica de
la gluconeogénesis
- En muchos tejidos, especialmente en el
corazón, los ácidos grasos son degradados
mediante un proceso conocido como
beta-oxidación, que produce acetil-CoA,
reingresado a su vuelta en el ciclo de Krebs
- La beta-oxidación también puede generar propionil-CoA, que
puede ser reingresado en la vía gluconeogénica hepática al
generar glucosa.
- La beta-oxidación también puede generar propionil-CoA, que
puede ser reingresado en la vía gluconeogénica hepática al
generar glucosa.
- En muchos tejidos, especialmente en el
corazón, los ácidos grasos son degradados
mediante un proceso conocido como
beta-oxidación, que produce acetil-CoA,
reingresado a su vuelta en el ciclo de Krebs
- En los organismos superiores,
el glicerol puede entrar en la
glucolisis a nivel hepático o ser
transformado en glucosa a
través de la hidroxiacetona
fosfato y el
gliceraldehído-3-fosfato,
siguiendo la ruta metabólica de
la gluconeogénesis
- En el catabolismo de los lípidos, los triglicéridos
son hidrolizados por enzimas lipasas para formar
ácidos grasos y glicero
- Cadena de transporte de electrones
- Una no tendría sentido sin la
otra en cuanto que el ATP y el
GTP producidos por el ciclo es
escaso y la producción de
NADH y FADH2 llevaría a un
entorno mitocondrial
excesivamente reducido
- mientras que la cadena respiratoria por sí sola necesitaría
una fuente de cofactores reducida para la oxidación del
entorno.
- Esta respiración celular extrae energía del NADH y FADH2, recreando
NAD+ y FAD y permitiendo de tal modo que el ciclo continue
- El ciclo de Krebs no usa oxígeno, que es utilizado en cambio en
la fosforilación oxidativa
- El ciclo de Krebs no usa oxígeno, que es utilizado en cambio en
la fosforilación oxidativa
- Esta respiración celular extrae energía del NADH y FADH2, recreando
NAD+ y FAD y permitiendo de tal modo que el ciclo continue
- mientras que la cadena respiratoria por sí sola necesitaría
una fuente de cofactores reducida para la oxidación del
entorno.
- Una no tendría sentido sin la
otra en cuanto que el ATP y el
GTP producidos por el ciclo es
escaso y la producción de
NADH y FADH2 llevaría a un
entorno mitocondrial
excesivamente reducido
- Catabolismo de los carbohidratos
Medienanhänge
{kind=link}
Möchten Sie kostenlos Ihre eigenen Mindmaps mit GoConqr erstellen? Mehr erfahren.