Beschreibung
|
|
Erstellt von Cher Bachar
vor mehr als 11 Jahre
|
|
Seite 1
{kind=link}
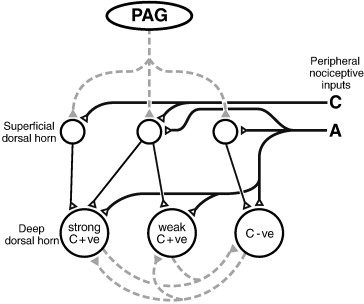
Fig. 2. A simplified model to explain how descending control from the PAG, which targets different populations of superficial dorsal horn neurons, could produce an inhibition of deep dorsal horn neurons that is proportional to their C-fiber input, but a facilitation of other neurons with weak or no C-fiber input. Solid lines represent direct (monosynaptic) connections, dotted lines represent indirect (polysynaptic) connections between neurons; open triangles/black lines are excitatory synapses and gray lines/filled triangles are inhibitory connections. The PAG inhibits superficial dorsal horn neurons that relay information carried by C-fibers to the deep dorsal horn. The net inhibitory or facilitatory effect of PAG stimulation is also a function of reciprocal inhibition between C+ve and C−ve neurons at the segmental level. C+ve: C-fiber input positive, C−ve: C-fiber input negative.
Background and methods- The aim of this study was to further understand the central processing of inputs arising from unmyelinated (C-fibres) and myelinated (A-fibres) nociceptors by (i) determining the response characteristics of Class 2 dorsal horn neurones to preferential activation of C- and A-fibre heat nociceptors, and (ii) investigating the control exerted by the dorsolateral/lateral region of the midbrain periaqueductal grey (DL/L-PAG) on C- and A-fibre-evoked responses of these neurones. The use of different rates of skin heating to preferentially activate unmyelinated (C-fibre; 2.5°C s-1) versus myelinated (A-fibre; 7.5°C s-1) heat nociceptors revealed that, Results- o in response to C-nociceptor activation, Class 2 neurones encode well only over the first 5°C above threshold, and that at higher temperatures responses decline. In contrast, responses to A-nociceptor activation are linear and encode skin temperature over more than 10°C, and almost certainly into the tissue-damaging range. o PAG stimulation raised thresholds and decreased significantly the magnitude of responses to A- and C-nociceptor activation. o However, differences were revealed in the effects of descending control on the relationships between skin temperature and neuronal firing rate; the linear relationship that occurred over the first 5°C of slow rates of skin heating was no longer evident, whereas that to fast rates of skin heating was maintained over the entire range, albeit shifted to the right. Interpretation- These data indicate that the sensori-discriminative information conveyed in A-fibre nociceptors is maintained and that the information from C-nociceptors is lost in the presence of descending control from the DL/L-PAG.
Background- Information about noxious events in the periphery is conveyed to the spinal cord in A- and C-fibre nociceptive afferents, which have largely distinct electrical and chemical properties and which convey different qualities of the pain signal. Descending control that originates in the different functional columns of the midbrain periaqueductal grey (PAG) has important roles in the modulation of spinal nociception in different behavioural and emotional states and, it is now believed, in animal models of chronic pain. However, few studies of descending control have considered differential modulation of A- versus C-nociceptor-evoked responses. Methods- stimulation of VL or DL/L PAG, coupled with noxious-pinch evoked responses of class 2 neurons (with C-fibre inputs) Results- pinch-evoked responses of class 2 neurons with C-fibre inputs were depressed, whereas those without C-fibre inputs were enhanced (probably A-mediated) Further experiments indicated these facilitatory effects were at least partly due to a reduction in C-fibre-mediated segmental inhibition. Interpretation- We suggest this differential control of spinal nociception would be appropriate in many of the varied situations in which the PAG is believed to become active, whether short term (e.g. fight or flight) or long term (e.g. chronic pain). Additionally, the pro-nociceptive effects observed in a subset of spinal neurons may be related to the descending facilitation that has been reported in animal models of chronic pain.
Taking together various evidence >> suggests that the differential effect of activating descending inhibition from the PAG on C-evoked activity in the deep dorsal horn is mediated by a direct post-synaptic action in the superficial dorsal horn. The strength of inhibitory control of the C-evoked activity in a given deep neuron reflects the degree to which that neuron receives C-fiber input relayed through the superficial layers. Because the strong C+ve deep dorsal horn neurons receive multiple inputs from the superficial dorsal horn, descending control produces a large net inhibition of nociceptive activity (both A- and C-fiber-evoked). By the same argument, activity of weak C+ve neurons, which receive a small input from the superficial dorsal horn, is only modestly depressed by PAG stimulation. Strong inhibitory influences on C+ve neurons will lift weak C+ve cells and C−ve cells from segmental inhibition, resulting in a net facilitatory effect. Importantly, direct A-nociceptor inputs to the deep dorsal horn that are not subject to descending control may act to further protect A-nociceptor-evoked responses. Together these mechanisms could account for the range in descending modulation of deep dorsal horn neurons, from strong inhibition to significant facilitation.
No C-fibre inputs
A few C-fibre inputs
Many C-fibre inputs
Mechanisms of differential descending control of C- versus A
McMullan and Lumb, 2006
Waters ad Lumb, 2008
Möchten Sie kostenlos Ihre eigenen Notizen mit GoConqr erstellen? Mehr erfahren.