13380929
Description
Mind Map by Laura Tafur, updated more than 1 year ago
|
|
Created by Laura Tafur
over 6 years ago
|
|
Distribución y transporte de proteínas
Annotations:
- Retículo endoplasmatico
- Retículo Endoplasmatico
- Es una red de túbulos y sacos (cisternas) que se extiende de la membrana nuclear por todo el citoplasma. Hay dos tipos distintos de RE que realizan diferentes funciones en la célula, estos son: RE rugoso y RE de transición
- El papel del retículo endoplasmatico se identificó como el lugar de síntesis de las proteínas destinadas a la secreción
- La mayoría de las proteínas son transferidas al RE mientras están siendo traducidas por los ribosomas unidos a la membrana. Por el contrario las proteínas destinas a permanecer en el citosol son sintetizadas en los ribosomas libres y liberadas al citosol cunado finaliza su traducción.
- El papel del retículo endoplasmatico se identificó como el lugar de síntesis de las proteínas destinadas a la secreción
- Marcaje de las proteínas para dirigirse al retículo endoplasmatico
- Las proteínas pueden ser translocadas al RE durante su síntesis en los ribosomas unidas a las membranas (translocación contraducional) o una vez que la traducción se ha completado en los ribosomas libres en el Citosol (translocación postraduccional).
- Translocación contraduccional
- Paso 1
- A medida que la secuencia señal emerge del ribosoma, es reconocida y unida a la particular de reconocimiento (PRS)
- A medida que la secuencia señal emerge del ribosoma, es reconocida y unida a la particular de reconocimiento (PRS)
- Paso 2
- La PRS acompaña al complejo hasta la membrana del RE, donde se une al receptor de la PRS.
- La PRS acompaña al complejo hasta la membrana del RE, donde se une al receptor de la PRS.
- Paso 3
- La PRS se libera, el ribosoma se une a un tranlocón y la secuencia señal se inserta en un canal de membrana.
- La PRS se libera, el ribosoma se une a un tranlocón y la secuencia señal se inserta en un canal de membrana.
- Paso 4
- La secuencia señal abre el translocón. se reanuda la traducción y la cadena polipeptídica en crecimiento es tranlocada a través de la membrana.
- La secuencia señal abre el translocón. se reanuda la traducción y la cadena polipeptídica en crecimiento es tranlocada a través de la membrana.
- Paso 5
- La escisión de la secuencia señal por la peptidasa de la señal libera el polipéptido en la luz del RE.
- La escisión de la secuencia señal por la peptidasa de la señal libera el polipéptido en la luz del RE.
- Paso 1
- Translocación postraduccional
- 1. Las proteínas destinadas a una internalización postraduccional al RE se sintetizan en ribosomas libres y se mantienen en una conformación desplegada mediante chaperonas citosólicas
- 2. sus secuencias señal son reconocidas por el complejo Sec62/63, que está asociado con el translocón en la membrana del RE.
- 3. La proteína Sec63 también está asociada con una proteína chaperona (BiP), que actúa como un <trinquete> molecular dirigiendo la translocación de la proteína al interior del RE.
- 1. Las proteínas destinadas a una internalización postraduccional al RE se sintetizan en ribosomas libres y se mantienen en una conformación desplegada mediante chaperonas citosólicas
- Translocación contraduccional
- Las proteínas pueden ser translocadas al RE durante su síntesis en los ribosomas unidas a las membranas (translocación contraducional) o una vez que la traducción se ha completado en los ribosomas libres en el Citosol (translocación postraduccional).
- Inserción de las proteínas en la membrana del RE
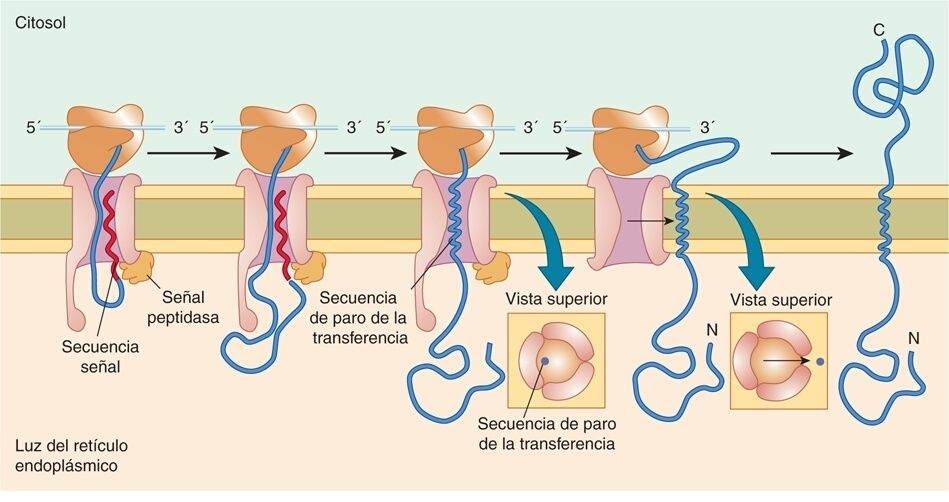
- Inserción de una proteína de membrana con una secuencia señal susceptible y una única secuencia de detención de la transferencia.
- 1. La secuencia de señal se escinde a medida que la cadena polipeptídica atraviesa la membrana.
- 2. La translocación de la cadena polipeptidica a través de la membrana es interrumpida.
- 3. El translocón se cierra y la proteína sale del canal lateralmente y se ancla en la membrana del RE.
- 4. La reanudación de la traducción da lugar a una proteína transmembrana.
- 1. La secuencia de señal se escinde a medida que la cadena polipeptídica atraviesa la membrana.
- Insercion de proteinas de membrana con secuencia señal internas que no se escinden
Annotations:
- insercion de proteinas de membrana con secuencia señal internas que no se escinden
- las secuencias señal internas que no se escinden pueden dar lugar a la inserción de cadenas polipeptídicas en cualquier orientacion en la membrana del RE
- Proceso
- A) Las secuencias señal dirige la insercion del polipéptido de tal forma que su extremo amino terminal queda expuesto al lado citosolico
- B) Otras secuencias señal internas se orientan de tal manera que dirigen la transferencia de la porcion amino terminal del polipéptido a través de la membrana. Al continuar la tradu ccion se forma una proteina que atraviesa la membrana la membrana del RE con su extremo amino terminal en la luz del RE
- A) Las secuencias señal dirige la insercion del polipéptido de tal forma que su extremo amino terminal queda expuesto al lado citosolico
- Proceso
- Inserción de una proteína de membrana con una secuencia señal susceptible y una única secuencia de detención de la transferencia.
- Plegamiento y procesamiento de las proteinas en el RE
- Con respecto a las proteinas que entran en la via secretora, muchos de estos acontecimeintos ocurren durante la translocacion a traves de la membrana del RE o en el interior de la luz del RE. Uno de estos procesos es la rotura proteolica del péptido secuencia señal a medida que la cadema polipeptidica se transloca a traves de la membrana dl RE.
- Las proteinas se translocan a traves de la membrana del RE a modo de cadenas polipeptidicas sin plegar mientras prosigue su traduccion, Por tanto estos polipeptidos se pliegan en su conformacion tridimensiones en el RE, asistidos, por chaperonas moleculares que facilitan el plegamiento de las cadenas polipeptidicas.
- la chaperona molecular BiP se une a las cadenas polipeptidicas cuando atraviesan la membrana del RE y favorece el plegamiento y el ensamble de proteinas en el RE
- la chaperona molecular BiP se une a las cadenas polipeptidicas cuando atraviesan la membrana del RE y favorece el plegamiento y el ensamble de proteinas en el RE
- Las proteinas se translocan a traves de la membrana del RE a modo de cadenas polipeptidicas sin plegar mientras prosigue su traduccion, Por tanto estos polipeptidos se pliegan en su conformacion tridimensiones en el RE, asistidos, por chaperonas moleculares que facilitan el plegamiento de las cadenas polipeptidicas.
- Con respecto a las proteinas que entran en la via secretora, muchos de estos acontecimeintos ocurren durante la translocacion a traves de la membrana del RE o en el interior de la luz del RE. Uno de estos procesos es la rotura proteolica del péptido secuencia señal a medida que la cadema polipeptidica se transloca a traves de la membrana dl RE.
- Control de calidad en el RE
- Muchas de las proteinas sintetizadas en el RE son rápidamente degradadas, principalmente porque no se pliegan correctamente; otras residen en el RE durante varias horas mientras que son plegadas correctamente. Asi un papel importante del RE es identificar las proteinas mal plegadas, marcadas y dirigirlas a una via de degradadcion
- El proceso de control de calidad del RE es complejo e implica BiP, otras chaperonas, la disulfuro proteina isomerasa, y un gran numero de proteinas accesorias
- En una vía conocida participan la glicoproteína chaperona, calreticulina, la cual reconoce los oligosacaridos sometidos a un procesamiento parcial de las glicoproteinas, recien traducidas y facilita el plegamiento correcto de la glicoproteina
- Plegamiento de proteinas por calreticulina
- 1, Durante su salida a traves del canal de translocon, se escinden dos residuos de glucosa de la glicoproteina para permitir la union de la calreticulina que facilitará el plegamiento
- 2. La escision del restante residuo de glucosa de la glucosa pone punto final a la interaccion con calreticulina y la glicoproteina para liberar a estas ultimas
- 3. Un sensor de plegamiento de proteinas valora las regiones hidrofobicas expuestas con el fin de evaluar el grado de plegamiento de la glicoproteina. La ausencia de estas regiones implica el plegamiento correcto de la proteina, que se dirige al RE transicional.
- 4.Si el plegamiento de la glicoproteina fuera incorrecto, el sensor, que posee actividad glucosiltransferasa añadira de nuevo un residuo de glucosa para que la glicoproteina regrese al ciclo de la chaperona carelticulina, Si el sensor detectara un número excesivamente alto de regiones hidrofobicas y la proteina no pudiera plegarse correctamente, se escindiran los residuos de manosa y se remitira de nuevo al citoplasma para su ubiquitinacion y degradacion por el proteasoma
- 1, Durante su salida a traves del canal de translocon, se escinden dos residuos de glucosa de la glicoproteina para permitir la union de la calreticulina que facilitará el plegamiento
- Respuesta a proteinas no plegadas
- La Proteina chaperona, BiP, participa en el plegamiento de las proteinas en la luz del RE
- A) En una celula no sometida a estres hay suficiente BiP disponible para plegar las proteinas recien sintetizadas y para mantener varios tipos de moleculas señalizadoras de la membrana del RE inactivas
- B) El estres celular, como el calor, el insulto quimico o la infeccion virica interfiere con el plegamiento proteico, de forma que se acumulan proteinas no plegadasno plegadas en el RE, de modo que estas son liberadas y se vuelven activas e inician la respuesra a proteinas no plegadas
- A) En una celula no sometida a estres hay suficiente BiP disponible para plegar las proteinas recien sintetizadas y para mantener varios tipos de moleculas señalizadoras de la membrana del RE inactivas
- La Proteina chaperona, BiP, participa en el plegamiento de las proteinas en la luz del RE
- En una vía conocida participan la glicoproteína chaperona, calreticulina, la cual reconoce los oligosacaridos sometidos a un procesamiento parcial de las glicoproteinas, recien traducidas y facilita el plegamiento correcto de la glicoproteina
- El proceso de control de calidad del RE es complejo e implica BiP, otras chaperonas, la disulfuro proteina isomerasa, y un gran numero de proteinas accesorias
- Muchas de las proteinas sintetizadas en el RE son rápidamente degradadas, principalmente porque no se pliegan correctamente; otras residen en el RE durante varias horas mientras que son plegadas correctamente. Asi un papel importante del RE es identificar las proteinas mal plegadas, marcadas y dirigirlas a una via de degradadcion
- RE Liso y sintesis de lipidos
- Ademas de su actividad en el procesamiento de las proteinas secretadas y de membrana, el RE es el sitio principal en el que se sintetizan los lipidos de la membrana en las celulas eucariotas. Puesto que son extremadamente hidrofobas, los lipidos se sintetizan asoaciados con membranas celulares ya existentes, en lugar de hacerlo en el ambiente acuoso del citosol.
- Las membranas de las celulas eucariotas estan compuestas por tres tipos fundamentales de lipidos: Fosfolipidos, glicolipidos y colesterol
- sintesis de fosfolipidos
- 1, Los glicerofosfolipidos se sintetizan en la membrana del RE a partir de precursores citosólicos. En primer lugar dos ácidos grasos unidos a los transportadores de coenzimas A ( coA) se unen al glicerol-3-fosfato, dando lugar al acido fosfatidico, que simultaneamente se inserta en la membrana.
- 2. A continucacion una fosfatasa convierte el acido fosfatídico en diacilglicerol. La union de diferentes grupos de cabeza polares al diacilglicerol. da lugar a la formacion de fosfatidilcolina, fosfadiletanolamina, fasfatidilserina y fofatidilinositol.
- 1, Los glicerofosfolipidos se sintetizan en la membrana del RE a partir de precursores citosólicos. En primer lugar dos ácidos grasos unidos a los transportadores de coenzimas A ( coA) se unen al glicerol-3-fosfato, dando lugar al acido fosfatidico, que simultaneamente se inserta en la membrana.
- Translocación de fosfolípidos através de la membrana del RE
- Puesto que los fosfolípidos se sintetizan en la cara citosólica de la membrana del RE, sólo se añaden a la mitad citosólica de la bicapa.
- Posteriormente se translocan a través de la membrana mediante flipasas fosfolipídicas, dando lugar a un crecimiento uniforme de las dos mitades de la bicapa fosfolipídica.
- Posteriormente se translocan a través de la membrana mediante flipasas fosfolipídicas, dando lugar a un crecimiento uniforme de las dos mitades de la bicapa fosfolipídica.
- Puesto que los fosfolípidos se sintetizan en la cara citosólica de la membrana del RE, sólo se añaden a la mitad citosólica de la bicapa.
- sintesis de fosfolipidos
- Las membranas de las celulas eucariotas estan compuestas por tres tipos fundamentales de lipidos: Fosfolipidos, glicolipidos y colesterol
- Ademas de su actividad en el procesamiento de las proteinas secretadas y de membrana, el RE es el sitio principal en el que se sintetizan los lipidos de la membrana en las celulas eucariotas. Puesto que son extremadamente hidrofobas, los lipidos se sintetizan asoaciados con membranas celulares ya existentes, en lugar de hacerlo en el ambiente acuoso del citosol.
- Exporacion de proteinas y lipidos desde el RE
- Tanto las proteinas como los lipidos migran atraves de la via secertora en vesiculkas de transporte, que se oroginan por gemacion en la membrana de un organulo y se funden posteriormente con la membrana de otro. Así, las moleculas son exportadas desde el RE y que en primer lugar, transportan su contenido al compartimiento intermedio RE-Golgi y despues al aparato de golgi
- Tanto las proteinas como los lipidos migran atraves de la via secertora en vesiculkas de transporte, que se oroginan por gemacion en la membrana de un organulo y se funden posteriormente con la membrana de otro. Así, las moleculas son exportadas desde el RE y que en primer lugar, transportan su contenido al compartimiento intermedio RE-Golgi y despues al aparato de golgi
- Exportación de proteínas y lípidos desde el RE
- Las moléculas son exportadas desde el RE en vesículas que se originan por gemación en el RE y que, en primer lugar, transportan su contenido al compartimento intermedio RE-Golgi y después al aparato de Golgi.
- Mientras la mayoría de las proteínas viajan desde el RE al Golgi, algunas proteínas han de retenerse en el RE en lugar de continuar a lo largo de la vía secretora. En cada caso, unas señales específicas de localización dirigen a las proteínas a sus destinos intracelulares correctos.
- La distinción entre las proteínas que han de ser exportadas y aquellas que son retenidas parece que se produce por dos tipos diferentes de secuencias que marcan a las proteínas.
- En el caso de la retención, hay presencia de una secuencia marcadora (KDEL) en su extremo carboxilo terminal, lo que obliga a las proteínas a ser retenidas en el RE. La retención de algunas proteínas transmembrana en el RE se caracteriza por una secuencia similar (KKXX).
- Dichas señales no impiden que las proteínas solubles del RE sean empaquetadas y transportadas al Golgi; sino que provocan que esas proteínas sean recuperadas a través de una vía de reciclado, siendo transportadas de vuelta al RE.
- Dichas señales no impiden que las proteínas solubles del RE sean empaquetadas y transportadas al Golgi; sino que provocan que esas proteínas sean recuperadas a través de una vía de reciclado, siendo transportadas de vuelta al RE.
- En el caso de la retención, hay presencia de una secuencia marcadora (KDEL) en su extremo carboxilo terminal, lo que obliga a las proteínas a ser retenidas en el RE. La retención de algunas proteínas transmembrana en el RE se caracteriza por una secuencia similar (KKXX).
- La distinción entre las proteínas que han de ser exportadas y aquellas que son retenidas parece que se produce por dos tipos diferentes de secuencias que marcan a las proteínas.
- Mientras la mayoría de las proteínas viajan desde el RE al Golgi, algunas proteínas han de retenerse en el RE en lugar de continuar a lo largo de la vía secretora. En cada caso, unas señales específicas de localización dirigen a las proteínas a sus destinos intracelulares correctos.
- Las moléculas son exportadas desde el RE en vesículas que se originan por gemación en el RE y que, en primer lugar, transportan su contenido al compartimento intermedio RE-Golgi y después al aparato de Golgi.
- Es una red de túbulos y sacos (cisternas) que se extiende de la membrana nuclear por todo el citoplasma. Hay dos tipos distintos de RE que realizan diferentes funciones en la célula, estos son: RE rugoso y RE de transición
- Aparato de Golgi
- Fábrica en la que proteínas recibidas del RE se procesan y distribuyen para ser transportadas a sus destinos finales. Además, sintetiza los glicolípidos y la esfingomielina.
- Organización del Golgi
- Morfológicamente, está compuesto por unas bolsas aplanadas, rodeadas de membrana (cisternas) y por vesículas asociadas.
- El AG cuenta con cierta polaridad tanto es su estructura como en su función, pues las proteínas provenientes del RE entran por su cara cis, que es convexa y se orienta al núcleo; y salen por su cara cóncava trans.
- El AG cuenta con cierta polaridad tanto es su estructura como en su función, pues las proteínas provenientes del RE entran por su cara cis, que es convexa y se orienta al núcleo; y salen por su cara cóncava trans.
- Se considera que el Golgi está constituido por cuatro regiones funcionalmente diferentes: Red cis del Golgi, apilamiento del Golgi (dividido en subcompartimentos medial y trans), y la red trans del Golgi. En ese orden:
- Las proteínas procedentes del RE se transportan al compartimento intermedio RE-Golgi y entran a continuación en el AG por la red cis del Golgi.
- Posteriormente pasan a los compartimentos medial y trans del apilamiento de Golgi, donde tienen lugar la mayor parte de las actividades metabólicas del AG.
- Las proteínas modificadas, los lípidos y los polisacáridos migran a continuación a la red trans del Golgi, que actúa como un centro de organización y distribución, dirigiendo el tráfico molecular a los lisosomas, a la membrana plasmática, o al exterior de la célula.
- Las proteínas modificadas, los lípidos y los polisacáridos migran a continuación a la red trans del Golgi, que actúa como un centro de organización y distribución, dirigiendo el tráfico molecular a los lisosomas, a la membrana plasmática, o al exterior de la célula.
- Posteriormente pasan a los compartimentos medial y trans del apilamiento de Golgi, donde tienen lugar la mayor parte de las actividades metabólicas del AG.
- Las proteínas procedentes del RE se transportan al compartimento intermedio RE-Golgi y entran a continuación en el AG por la red cis del Golgi.
- Morfológicamente, está compuesto por unas bolsas aplanadas, rodeadas de membrana (cisternas) y por vesículas asociadas.
- Glicosilación de proteínas en el Golgi
- El procesamiento de las proteínas en el Golgi supone la modificación y síntesis de los restos de carbohidratos de las glicoproteínas.
- Las proteínas se modifican en el RE al añadírseles un oligosacárido constituido por 14 residuos de azúcar. Luego, son eliminados 3 residuos de glucosa y 1 manosa, mientras lo polipéptidos siguen el RE.
- Los N-oligosacáridos se procesan en el AG por una secuencia ordenada de reacciones. La primera modificación de las proteínas destinadas a ser secretadas o de la membrana plasmática, es la eliminación de otros 3 residuos de manosa.
- Seguido, la adición secuencial de una N-acetilglucosamina, la eliminación de 2 manosas más, y la adición de una fucosa y de otras 2 N-acetilglucosaminas.
- Seguido, la adición secuencial de una N-acetilglucosamina, la eliminación de 2 manosas más, y la adición de una fucosa y de otras 2 N-acetilglucosaminas.
- El procesamiento de N-oligosacárido de las proteínas lisosómicas difiere del de las proteínas secretadas y de la membrana plasmática. Las destinadas a incorporarse en los lisosomas, en vez de la eliminación inicial de tres residuos de manosa, son modificadas mediante una fosforilación de la manosa.
- Los N-oligosacáridos se procesan en el AG por una secuencia ordenada de reacciones. La primera modificación de las proteínas destinadas a ser secretadas o de la membrana plasmática, es la eliminación de otros 3 residuos de manosa.
- Las proteínas se modifican en el RE al añadírseles un oligosacárido constituido por 14 residuos de azúcar. Luego, son eliminados 3 residuos de glucosa y 1 manosa, mientras lo polipéptidos siguen el RE.
- El procesamiento de las proteínas en el Golgi supone la modificación y síntesis de los restos de carbohidratos de las glicoproteínas.
- Metabolismo de lípidos y polisacáridos en el Golgi
- El AG participa en el metabolismo lipídico (concretamente, en las síntesis de glicolípidos y esfingomielina). La esfingomielina es sintetizada por la transferencia de un grupo fosforilcolina desde la fosfatidilcolina a la ceramida. Alternativamente, la adición de carbohidratos a la ceramida puede dar lugar a diferentes glicolípidos.
- En las células vegetales, el AG tiene la función añadida de ser el lugar donde se sintetizan los polisacáridos complejos de la pared celular.
- El AG participa en el metabolismo lipídico (concretamente, en las síntesis de glicolípidos y esfingomielina). La esfingomielina es sintetizada por la transferencia de un grupo fosforilcolina desde la fosfatidilcolina a la ceramida. Alternativamente, la adición de carbohidratos a la ceramida puede dar lugar a diferentes glicolípidos.
- Distribución y exportación de proteínas desde el AG
- Las proteínas son transportadas desde el AG a sus destinos finales a través de la vía secretora. Esto implica que las proteínas se distribuyan en diferentes tipos de vesículas de transporte, las cuales saldrán por gemación desde la red trans y llevarán su contenido hasta la localización celular adecuada.
- Algunas proteínas se transportan desde el Golgi a la membrana plasmática por una vía secretora constitutiva, responsable de la incorporación de nuevas proteínas y lípidos a la membrana plasmática.
- Otras proteínas son transportadas a la superficie celular a través de una vía diferente de secreción regulada, o son dirigidas específicamente a otros destinos intracelulares, como los lisosomas o las vacuolas.
- La vía secretora constitutiva, que está presente en todas las células, da lugar a una secreción de proteínas continua no regulada. Sin embargo, algunas células también poseen una vía secretora regulada diferente, a través de la que secretan proteínas específicas en respuesta a señales ambientales.
- Las proteínas son transportadas desde el AG a sus destinos finales a través de la vía secretora. Esto implica que las proteínas se distribuyan en diferentes tipos de vesículas de transporte, las cuales saldrán por gemación desde la red trans y llevarán su contenido hasta la localización celular adecuada.
- Fábrica en la que proteínas recibidas del RE se procesan y distribuyen para ser transportadas a sus destinos finales. Además, sintetiza los glicolípidos y la esfingomielina.
- Mecanismo de transporte de las vesículas
- El transporte de las vesículas es una actividad celular fundamental, responsable del tráfico molecular entre diversos compartimentos rodeados por membrana, específicos. Por lo tanto, la selectividad de dicho transporte resulta clave para mantener la organización funcional de la célula.
- Aproximaciones experimentales al conocimiento del transporte de las vesículas
- Tres abordajes experimentales han permitido avanzar en el conocimiento de los mecanismos del transporte de las vesículas.
- El aislamiento de mutantes de levaduras defectuosas en el transporte y distribución de las proteínas.
- El análisis bioquímico de las vesículas sinápticas, que son las responsables de la secreción regulada de los neurotransmisores por las neuronas.
- La reconstitución del transporte de vesículas en sistemas acelulares.
- Lo más importante de estos experimentos, es el hecho de que los resultados obtenidos han convergido, indicando que la secreción está regulada por mecanismos moleculares similares en células distintas entre sí.
- El aislamiento de mutantes de levaduras defectuosas en el transporte y distribución de las proteínas.
- Tres abordajes experimentales han permitido avanzar en el conocimiento de los mecanismos del transporte de las vesículas.
- El transporte de las vesículas es una actividad celular fundamental, responsable del tráfico molecular entre diversos compartimentos rodeados por membrana, específicos. Por lo tanto, la selectividad de dicho transporte resulta clave para mantener la organización funcional de la célula.
Media attachments
{kind=link}
Want to create your own Mind Maps for free with GoConqr? Learn more.