20948665
Description
Mind Map by Jacces Luna, updated more than 1 year ago
|
|
Created by Jacces Luna
almost 5 years ago
|
|
CAPACIDAD DE DIFUSION
Y CONTROL DE VENTILADOS
- Mediciones de gases en sangre
- La medición del contenido de
oxígeno de la sangre (en mililitros
de O2 por 100 ml de sangre) es
un proceso laborioso
- Afortunadamente se ha creado un
electrodo de oxígeno que produce una
corriente eléctrica en proporción a la
concentración de oxígeno disuelto. Si
este electrodo se coloca en un líquido
mientras se hace burbujear oxígeno
artificialmente hacia él, la corriente
producida por el electrodo de oxígeno
aumentará hasta un valor máximo.
- A este valor máximo, el líquido está
saturado con oxígeno es decir, todo el
oxígeno que puede disolverse a esa
temperatura y PO2 está disuelto. A
una temperatura constante, la
cantidad disuelta y, así, la corriente
eléctrica, sólo dependen de la PO2 del
gas.
- Por conveniencia, ahora puede
decirse que el líquido tiene la misma
PO2 que el gas.
- La cantidad real de oxígeno disuelto en estas
circunstancias no es en particular importante
(puede consultarse en cuadros de solubilidad, si
se desea); simplemente es una función lineal de
la PO2.
- Una PO2 más baja
indica que hay menos
oxígeno disuelto; una
PO2 más alta indica
que hay más oxígeno
disuelto.
- Si el electrodo de oxígeno
se inserta a continuación
en una muestra
desconocida de sangre, la
PO2 de esa muestra puede
leerse de manera directa a
partir de la escala
previamente calibrada.
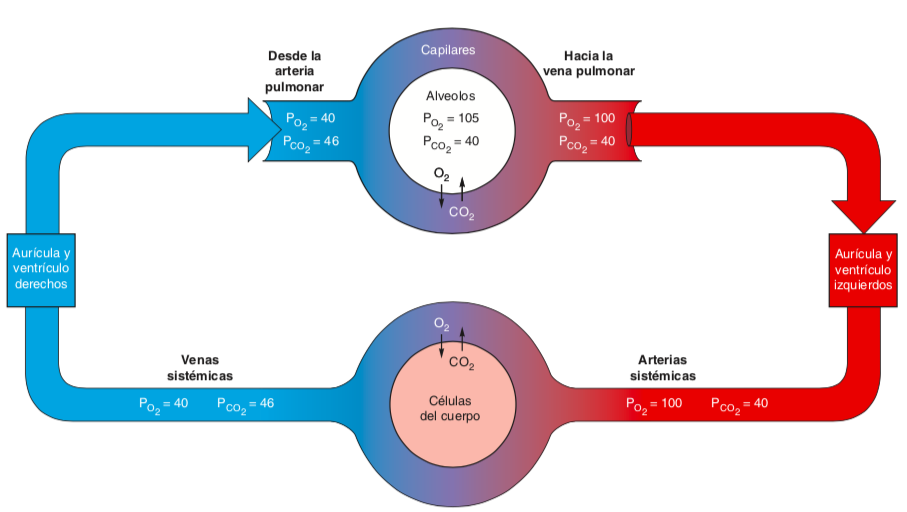
- El aire alveolar tiene una PO2
de alrededor de 105 mm Hg,
de modo que esta lectura
indica que la sangre está en
equilibrio casi completo con el
aire alveolar
- El aire alveolar tiene una PO2 de
alrededor de 105 mm Hg, de modo
que esta lectura indica que la sangre
está en equilibrio casi completo con
el aire alveolar.
- El electrodo de oxígeno sólo responde
al oxígeno disuelto en agua o plasma;
no puede mostrar respuesta al oxígeno
que está unido a la hemoglobina en los
eritrocitos.
- El electrodo de oxígeno sólo responde
al oxígeno disuelto en agua o plasma;
no puede mostrar respuesta al oxígeno
que está unido a la hemoglobina en los
eritrocitos.
- El aire alveolar tiene una PO2 de
alrededor de 105 mm Hg, de modo
que esta lectura indica que la sangre
está en equilibrio casi completo con
el aire alveolar.
- El aire alveolar tiene una PO2
de alrededor de 105 mm Hg,
de modo que esta lectura
indica que la sangre está en
equilibrio casi completo con el
aire alveolar
- Si el electrodo de oxígeno
se inserta a continuación
en una muestra
desconocida de sangre, la
PO2 de esa muestra puede
leerse de manera directa a
partir de la escala
previamente calibrada.
- Una PO2 más baja
indica que hay menos
oxígeno disuelto; una
PO2 más alta indica
que hay más oxígeno
disuelto.
- La cantidad real de oxígeno disuelto en estas
circunstancias no es en particular importante
(puede consultarse en cuadros de solubilidad, si
se desea); simplemente es una función lineal de
la PO2.
- Por conveniencia, ahora puede
decirse que el líquido tiene la misma
PO2 que el gas.
- A este valor máximo, el líquido está
saturado con oxígeno es decir, todo el
oxígeno que puede disolverse a esa
temperatura y PO2 está disuelto. A
una temperatura constante, la
cantidad disuelta y, así, la corriente
eléctrica, sólo dependen de la PO2 del
gas.
- Afortunadamente se ha creado un
electrodo de oxígeno que produce una
corriente eléctrica en proporción a la
concentración de oxígeno disuelto. Si
este electrodo se coloca en un líquido
mientras se hace burbujear oxígeno
artificialmente hacia él, la corriente
producida por el electrodo de oxígeno
aumentará hasta un valor máximo.
- La medición del contenido de
oxígeno de la sangre (en mililitros
de O2 por 100 ml de sangre) es
un proceso laborioso
- Importancia de las
mediciones de la PO2 y
la PCO2 en la sangre
- Puesto que el oxígeno en los
eritrocitos no afecta de manera
directa las mediciones de la PO2
en sangre, la PO2 no proporciona
una medición del contenido total
de oxígeno de la sangre entera.
- Aun así,
proporciona un
buen índice de la
función pulmonar.
- Por ejemplo, si el aire inspirado
tiene una PO2 normal, pero la PO2
arterial está por debajo de lo
normal, se concluiría que el
intercambio de gases en los
pulmones está alterado.
- Así, las mediciones de la PO2 arterial
proporcionan información valiosa en el
tratamiento de personas que tienen
enfermedades pulmonares, en la práctica
de intervenciones quirúrgicas (cuando la
respiración puede estar deprimida por la
anestesia), y en el cuidado de lactantes
prematuros que presentan síndrome de
dificultad respiratoria.
- Cuando los pulmones están
funcionando de manera
apropiada, la PO2 de la sangre
arterial sistémica es de sólo 5
mm Hg menos que la PO2 del
aire alveolar.
- A una PO2 normal de alrededor de 100 mm
Hg, dor de 100 mm Hg, la hemoglobina está
cargada casi por completo con oxígeno, lo
cual es indicado por una saturación de
oxihemoglobina (el porcentaje de
oxihemoglobina de la hemoglobina total) de
97%.
- A una PO2 normal de alrededor de 100 mm
Hg, dor de 100 mm Hg, la hemoglobina está
cargada casi por completo con oxígeno, lo
cual es indicado por una saturación de
oxihemoglobina (el porcentaje de
oxihemoglobina de la hemoglobina total) de
97%.
- Cuando los pulmones están
funcionando de manera
apropiada, la PO2 de la sangre
arterial sistémica es de sólo 5
mm Hg menos que la PO2 del
aire alveolar.
- Así, las mediciones de la PO2 arterial
proporcionan información valiosa en el
tratamiento de personas que tienen
enfermedades pulmonares, en la práctica
de intervenciones quirúrgicas (cuando la
respiración puede estar deprimida por la
anestesia), y en el cuidado de lactantes
prematuros que presentan síndrome de
dificultad respiratoria.
- Por ejemplo, si el aire inspirado
tiene una PO2 normal, pero la PO2
arterial está por debajo de lo
normal, se concluiría que el
intercambio de gases en los
pulmones está alterado.
- Aun así,
proporciona un
buen índice de la
función pulmonar.
- Puesto que el oxígeno en los
eritrocitos no afecta de manera
directa las mediciones de la PO2
en sangre, la PO2 no proporciona
una medición del contenido total
de oxígeno de la sangre entera.
- REGULACIÓN
DE LA
RESPIRACIÓN
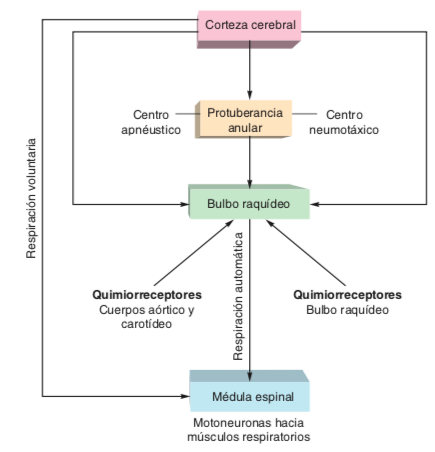
- Las neuronas motoras que estimulan los
músculos respiratorios están controladas por
dos vías descendentes principales: una que
controla la respiración voluntaria, y otra que
controla la respiración involuntaria
- El control rítmico inconsciente de la
respiración está influido por retroacción
sensorial proveniente de receptores
sensibles a la PCO2, el pH y la PO2 de la
sangre arterial.
- La inspiración y espiración se producen por la
contracción y relajación de músculos
esqueléticos en respuesta a la actividad en
neuronas motoras somáticas en la médula
espinal. La actividad de estas neuronas
motoras está controlada, a su vez, por tractos
descendentes provenientes de neuronas en
los centros de control respiratorio en el bulbo
raquídeo, y de neuronas en la corteza cerebral.
- Centros respiratorios del tallo
encefálico
- Las neuronas motoras somáticas que
estimulan los músculos respiratorios
tienen su cuerpo celular en la sustancia
gris de la médula espinal.
- Las motoneuronas del nervio frénico, que
estimulan el diafragma, tienen cuerpos
celulares en el nivel cervical de la médula
espinal; las que inervan los músculos
respiratorios de la caja torácica y el
abdomen tienen cuerpos celulares en la
región toracolumbar de la médula espinal.
- El ritmo respiratorio es generado por
una agregación laxa de neuronas en
la región ventrolateral del bulbo
raquídeo, que forma el centro de la
ritmicidad para el control de la
respiración automática.
- Quimiorreceptores
- El control automático de la
respiración también está influido por
aferencias provenientes de
quimiorreceptores, que son en
conjunto sensibles a cambios del pH
del líquido intersticial del encéfalo y
del líquido cefalorraquídeo, y de la
PCO2, el pH y la PO2 de la sangre.
- Hay dos grupos de
quimiorreceptores que muestran
respuesta a cambios de la PCO2, el
pH y la PO2; se trata de los
quimiorreceptores centrales en el
bulbo raquídeo, y los
quimiorreceptores periféricos.
- Los quimiorreceptores periféricos incluyen
los cuerpos aórticos, ubicados alrededor del
arco aórtico, y los cuerpos carotídeos,
situados en cada arteria carótida primitiva
en el punto donde se ramifican hacia las
arterias carótidas interna y externa
- Quimiorreceptores en
el bulbo raquídeo
- Los quimiorreceptores más sensibles a cambios de la PCO2
arterial están situados en la superficie ventrolateral del bulbo
raquídeo, cerca de la salida de los pares craneales IX y X.
Estas neuronas quimiorreceptoras están separadas desde el
punto de vista anatómico de las neuronas del centro de la
ritmicidad en el bulbo raquídeo, pero se comunican mediante
sinapsis con ellas.
- Quimiorreceptores
periféricos
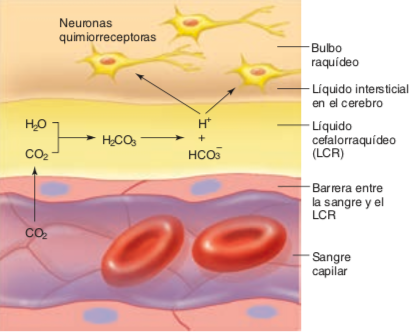
- El CO2 en sangre no estimula de
manera directa los cuerpos aórtico y
carotídeo.
- En lugar de eso, son estimulados por un aumento
de la concentración de H+ (disminución del pH) de
la sangre arterial, que ocurre cuando aumenta el
CO2 y, así, el ácido carbónico, en sangre.
- En lugar de eso, son estimulados por un aumento
de la concentración de H+ (disminución del pH) de
la sangre arterial, que ocurre cuando aumenta el
CO2 y, así, el ácido carbónico, en sangre.
- El CO2 en sangre no estimula de
manera directa los cuerpos aórtico y
carotídeo.
- Quimiorreceptores
periféricos
- Los quimiorreceptores más sensibles a cambios de la PCO2
arterial están situados en la superficie ventrolateral del bulbo
raquídeo, cerca de la salida de los pares craneales IX y X.
Estas neuronas quimiorreceptoras están separadas desde el
punto de vista anatómico de las neuronas del centro de la
ritmicidad en el bulbo raquídeo, pero se comunican mediante
sinapsis con ellas.
- Quimiorreceptores en
el bulbo raquídeo
- Los quimiorreceptores periféricos incluyen
los cuerpos aórticos, ubicados alrededor del
arco aórtico, y los cuerpos carotídeos,
situados en cada arteria carótida primitiva
en el punto donde se ramifican hacia las
arterias carótidas interna y externa
- Hay dos grupos de
quimiorreceptores que muestran
respuesta a cambios de la PCO2, el
pH y la PO2; se trata de los
quimiorreceptores centrales en el
bulbo raquídeo, y los
quimiorreceptores periféricos.
- El control automático de la
respiración también está influido por
aferencias provenientes de
quimiorreceptores, que son en
conjunto sensibles a cambios del pH
del líquido intersticial del encéfalo y
del líquido cefalorraquídeo, y de la
PCO2, el pH y la PO2 de la sangre.
- Quimiorreceptores
- El ritmo respiratorio es generado por
una agregación laxa de neuronas en
la región ventrolateral del bulbo
raquídeo, que forma el centro de la
ritmicidad para el control de la
respiración automática.
- Las motoneuronas del nervio frénico, que
estimulan el diafragma, tienen cuerpos
celulares en el nivel cervical de la médula
espinal; las que inervan los músculos
respiratorios de la caja torácica y el
abdomen tienen cuerpos celulares en la
región toracolumbar de la médula espinal.
- Las neuronas motoras somáticas que
estimulan los músculos respiratorios
tienen su cuerpo celular en la sustancia
gris de la médula espinal.
- Centros respiratorios del tallo
encefálico
- La inspiración y espiración se producen por la
contracción y relajación de músculos
esqueléticos en respuesta a la actividad en
neuronas motoras somáticas en la médula
espinal. La actividad de estas neuronas
motoras está controlada, a su vez, por tractos
descendentes provenientes de neuronas en
los centros de control respiratorio en el bulbo
raquídeo, y de neuronas en la corteza cerebral.
- El control rítmico inconsciente de la
respiración está influido por retroacción
sensorial proveniente de receptores
sensibles a la PCO2, el pH y la PO2 de la
sangre arterial.
- Las neuronas motoras que estimulan los
músculos respiratorios están controladas por
dos vías descendentes principales: una que
controla la respiración voluntaria, y otra que
controla la respiración involuntaria
Media attachments
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Want to create your own Mind Maps for free with GoConqr? Learn more.