
Description
|
|
Created by Emily Rowland-Rawson
over 6 years ago
|
|
Page 1
9.1 Transport systems in dicotyledonous plants
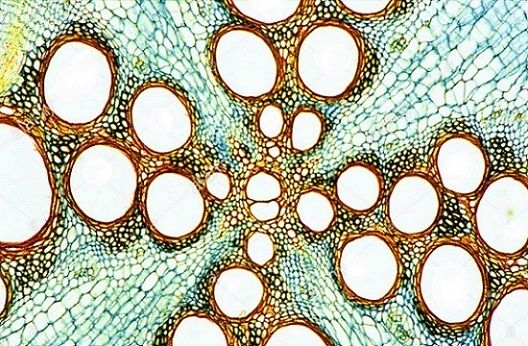
Dicotyledonous plants (dicots) make seeds that contain two cotyledons, organs that act as food stores for the developing embryo plant and form the first leaves when the seed germinates. There are herbaceous dicots, with soft tissues and a relatively short life cycle (leaves and stems that die down at the end of the growing season to the soil level), and woody (arborescent) dicots, which have hard, lignified tissues and a long life cycle (in some cases hundreds of years). Dicotyledons have a series of transport vessels running through the stem, roots, and leaves. This is known as the vascular system. In herbaceous dicots this is made up of two main types of transport vessels, the xylem and the phloem, described later in this topic. These transport tissues are arranged together in vascular bundles in the leaves, stems, and roots of herbaceous dicots. The picture below shows the vascular bundle in a root.
{kind=link}
Structure and functions of xylem Xylem is a mainly dead tissue that has two main functions: the transport of water and mineral ions, and structural support. Xylem contains four types of cell: Vessels: consist of dead, hollow cells with wide lumens. Cells are linked end to end in a drainpipe fashion. End walls have one or more perforations - allows rapid transport of large volumes of water from roots up through the stem. Tracheids: dead, hollow cells with narrower lumens than vessels. Tracheids are connected vertically to each other via bordered pits - Conduct water in conifers which, because they have needles, do not lose as much water as, for example, broadleaved trees. Narrower lumens encourages capillarity. Tracheids are elongated cells in the xylem of vascular plants that serve in the transport of water and mineral salts. Tracheids are one of two types of tracheary elements, vessel elements being the other. Tracheids, unlike vessel elements, do not have perforation plates. Parenchyma: Thin cellulose walls with living contents - contribute to support via turgidity. All parenchyma cells are living at maturity and capable of cell division. Fibres: Similar to fibres in sclerenchyma - provide strength. They are non-living sclerenchyma cells as they lose their protoplast at maturity.
The two main groups in which secondary xylem can be found are: Gymnosperms (conifers) have only tracheids for water transport. there are some six hundred species of conifers. All species have secondary xylem, which is relatively uniform in structure throughout this group. Many conifers become tall trees: the secondary xylem of such trees is used and marketed as softwood. angiosperms (flowering plants) have both vessels and tracheids, and primarily rely on vessels for water transport.there are some quarter of a million to four hundred thousand species of angiosperms. Within this group secondary xylem is rare in the monocots. Many non-monocot angiosperms become trees, and the secondary xylem of these is used and marketed as hardwood. In Conifers, the xylem, or water-conducting tissue, is made up exclusively of tracheids. The gymnosperms and angiosperms together compose the spermatophytes, or seed plants.
Structure and functions of phloem In vascular plants, phloem is the living tissue that transports the soluble organic compounds made during photosynthesis and known as photosynthates, in particular the sugar sucrose, to parts of the plant where needed. This transport process is called translocation. In trees, the phloem is the innermost layer of the bark, hence the name, derived from the Greek word φλοιός (phloios) meaning "bark".
Wider reading: Non-vascular plants and Bryophytes Non-vascular plants are plants without a vascular system consisting of xylem and phloem. Although non-vascular plants lack these particular tissues, many possess simpler tissues that are specialized for internal transport of water. Bryophytes are an informal group consisting of three divisions of non-vascular land plants (embryophytes): the liverworts, hornworts and mosses.They are characteristically limited in size and prefer moist habitats although they can survive in drier environments. The bryophytes consist of about 20,000 plant species. Bryophytes produce enclosed reproductive structures (gametangia and sporangia), but they do not produce flowers or seeds. They reproduce via spores. Bryophytes are usually considered to be a paraphyletic group and not a monophyletic group, although some studies have produced contrary results. The term "bryophyte" comes from Greek βρύον, bryon "tree-moss, oyster-green" and φυτόν, phyton "plant". The defining features of bryophytes are: Their life cycles are dominated by the gametophyte stage Their sporophytes are unbranched They do not have a true vascular tissue containing lignin (although some have specialized tissues for transporting water) Because these plants lack lignified water-conducting tissues, they can't become as tall as most vascular plants.
Page 2
9.2 Water transport in multicellular plants
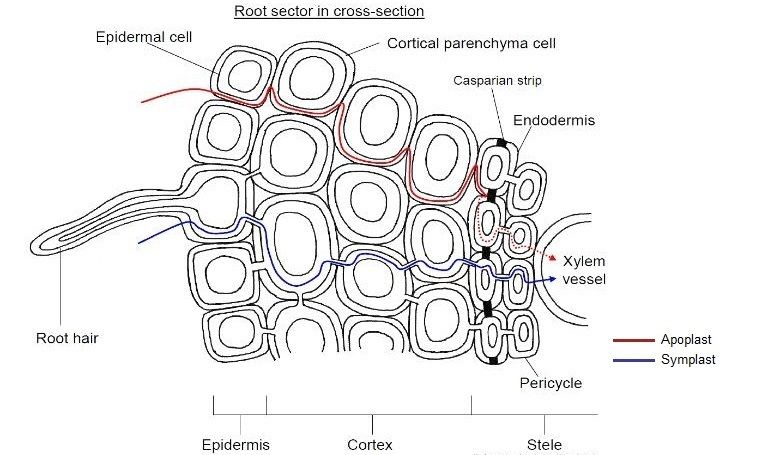
Soil water has a very low concentration of dissolved minerals so it has a very high water potential. The cytoplasm and vacuolar sap of the root hair cell (and the other root cells) contain many different solvents including sugars, mineral ions, and amino acids so the water potential in the cell is lower. As a result water moves into the root hair cells by osmosis. Once the water has moved into the root hair cell it continues to move across the root to the xylem in one of two different pathways, apoplast and symplast.
The need for water in plants Obviously, plants need water to live because they die if they do not have water for an extended period of time. However this is not solely due to water being used for photosynthesis or mineral transport. Water also provides structural support that allows plants to stand upright. Hydrostatic pressure (or turgor pressure) as a result of osmosis in plant cells provides a hydrostatic skeleton to support the stems and leaves. So, for example, the turgor pressure in leaf cells is around 1.5 M.Pa- that is 11251mmHg (unit of pressure still commonly used in medicine) human systolic blood pressure is around 120mmHg. That's pretty incredible for a plant. Turgor also drives cell expansion- it is the force that enables plant roots to force their way through tarmac and concrete. Transpiration (the loss of water by evaporation) helps to keep plants cool. Mineral ions and the products of photosynthesis are transported in aqueous solutions. Water is needed for photosynthesis.
Root hair adaptations Root hair cells are the exchange surface in plants where water is taken into the body of the plant from the soil. A root hair is a long, thin extension from a root hair cell, a specialised epidermal cell found near the growing root tip. Here are some adaptations of root hairs that make them efficient exchange surfaces. Their microscopic size means they can penetrate easily between soil particles. Each microscopic hair has a large SA: V ratio and there are thousands on each growing root tip. Each hair has a thin surface layer (just the cell wall and cell-surface membrane) through which diffusion and osmosis can rake place quickly. The concentration of solutes in the cytoplasm of root hair cells maintains a water potential gradient between the soil water and the cell.
{kind=link}
Apoplast pathway This is the movement of water through the apoplast- the cell walls and the inter-cellular spaces. Water fills the spaces between the open network of fibres in the cellulose cell wall. As water molecules move into the xylem, more water molecules are pulled through the apoplast behind them due to the cohesive forces between the water molecules. The pull from water moving into the xylem and up the plant along with the cohesive forces between the water molecules creates a tension that means there is a continuous flow of water through the open structure of the cellulose wall, which offers little or no resistance. The term apoplast was coined in 1930 by Münch in order to separate the "living" symplast from the "dead" apoplast.
Symplast pathway Water moves through the symplast- the continuous cytoplasm of the living plant cells that is connected through the plasmodesmata - by osmosis. The root hair cell has a higher water potential than the next cell along. This is the result of water diffusing in from the soil which has made the cytoplasm more dilute. So water moves from the root hair cell into the next door cell by osmosis. This process continues from cell to cell across the root until the xylem is reached. As water leaves the root hair cell by osmosis, the water potential of the cytoplasm falls again, and this maintains a steep water potential gradient to ensure that as much water as possible continues to move into the cell from the soil.
Movement of water into the xylem Water moves across the root in the apoplast and symplast pathways until it reaches the endodermis- the layer of cells surrounding the vascular tissue (xylem and phloem) of the roots. The endoderm is is particularly noticeable in the roots because of the effect of the Casparian strip. The Casparian strip is a band of waxy material called suberin that runs around each of the endodermal cells forming a waterproof layer. At this point, water in the apoplast pathway can go no further and it is forced into the cytoplasm of the cell, joining the water in the symplast pathway. Th is diversion to the cytoplasm is significant as to get to get there, water must pass through the selectively permeable cell surface membranes, this excludes any potentially-toxic solutes in the soil water from reaching living tissues, as the membranes would have no carrier proteins to admit them. Once forced into the cytoplasm the water joins the symplast pathway. The solute concentration in the cytoplasm of the endodermal cells is relatively dilute compared to the cells in the xylem. In addition, it appears that the endodermal cells move mineral ions into the xylem by active transport. As a result the water potential of the xylem cells is much lower than the water potential of the endodermal cells. This increases the rate of water moving into the xylem by osmosis down a water potential gradient from the endodermis through the symplast pathway. Once inside the vascular bundle, water returns to the apoplast pathway to enter the xylem itself and move up the plant. The active pumping of minerals into the xylem to produce movement of water by osmosis results in root pressure and it is independent of any effects of transpiration. Root pressure gives water a push up the xylem, but under most circumstances it is not the major factor in the movement of water up from the roots to the leaves.
Evidence for the involvement of active transport in root pressure It has taken some time and many different investigations to determine the role of active transport in moving water from the root endodermis into the xylem. There are several strands of evidence supporting the current model: Some poisons, such as cyanide, affect the mitochondria and prevent the production of ATP. If cyanide is applied to root cells so there is no energy supply, the root pressure disappears. This suggests that energy is needed to maintain root pressure. Root pressure increases with a rise in temperature and falls with a fall in temperature, suggesting this is a result of a chemical reaction. If levels of oxygen or respiratory substrates fall, root pressure falls. This suggests that respiration is involved Xylem sap may exude from the cut end of stems at certain times. In the natural world, xylem sap is forced out of special pores at the ends of leaves in some conditions - for example overnight, when transpiration is low. This is known as guttation.
Key terms water potential: a measure of the quantity of water compared to solutes, measured as the pressure created by the water molecules in kilopascals (kPa), can be written ψ (psi). The potential of pure water (Ψwpure H2O) is designated a value of zero (even though it isn't) osmosis: the diffusion of water down a water potential gradient through a partially permeable membrane (a passive process) active transport: the movement of particles across a plasma membrane against a concentration gradient (a process requiring energy) plasmodesmata: a thin strand of cytoplasm that passes through the cell walls of adjacent plant cells
Page 3
9.3 Transpiration
Photosynthesis, the process by which green plants make their own food, takes place mainly in the leaves. Carbon dioxide and water are both needed so, for successful photosynthesis to take place in a leaf, water must be transported there from the roots and carbon dioxide must be taken into the cells of the leaf from the air. Carbon dioxide diffuses into the leaf cells down a concentration gradient from the air spaces within the leaf. In a process of gaseous exchange, oxygen also moves out of the leaf cells into the air spaces by diffusion down a concentration gradient (oxygen is a waste product of photosynthesis). At the same time water evaporates from the surfaces of the leaf cells in to the air spaces.
Effects on transpiration Number of leaves: More leaves (or spines, or other photosynthesizing organs) means a bigger surface area and more stomata for gaseous exchange. This will result in greater water loss. Number of stomata: More stomata will provide more pores for transpiration. Size of the leaf: A leaf with a bigger surface area will transpire faster than a leaf with a smaller surface area. Presence of plant cuticle: A waxy cuticle is relatively impermeable to water and water vapour and reduces evaporation from the plant surface except via the stomata. A reflective cuticle will reduce solar heating and temperature rise of the leaf, helping to reduce the rate of evaporation. Tiny hair-like structures called trichomes on the surface of leaves also can inhibit water loss by creating a high humidity environment at the surface of leaves. These are some examples of the adaptations of plants for conservation of water that may be found on many xerophytes. Light supply: The rate of transpiration is controlled by stomatal aperture, and these small pores open especially for photosynthesis. While there are exceptions to this (such as night or "CAM photosynthesis"), in general a light supply will encourage open stomata. Temperature: Temperature affects the rate in two ways; An increased rate of evaporation due to a temperature rise will hasten the loss of water. And decreased relative humidity outside the leaf will increase the water potential gradient. Relative humidity: Drier surroundings gives a steeper water potential gradient, and so increases the rate of transpiration. Wind: In still air, water lost due to transpiration can accumulate in the form of vapor close to the leaf surface. This will reduce the rate of water loss, as the water potential gradient from inside to outside of the leaf is then slightly less. Wind blows away much of this water vapor near the leaf surface, making the potential gradient steeper and speeding up the diffusion of water molecules into the surrounding air. Even in wind, though, there may be some accumulation of water vapor in a thin boundary layer of slower moving air next to the leaf surface. The stronger the wind, the thinner this layer will tend to be, and the steeper the water potential gradient. Water supply: Water stress caused by restricted water supply from the soil may result in stomatal closure and reduce the rates of transpiration.
Page 4
9.4 Translocation
The leaves of a plant produce large amounts of glucose, which is needed for respiration by all the cells of the plant. The glucose is convened to sucrose for transport. When it reaches the cells where it is needed it is converted back to glucose for respiration, or to starch for storage, or used to produce the amino acids and other compounds needed within the cell.
Page 5
9.5 Plant adaptations to water availability
Land plants exist in a state of constant compromise between getting the carbon dioxide they need for photosynthesis and losing the water they need for turgor pressure and transport. They must have a large SA:V ratio for gaseous exchange and the capture of light for photosynthesis, but this greatly increases their risk of water loss by transpiration.
0 comments
Want to create your own Notes for free with GoConqr? Learn more.