26806528
MATRIZ EXTRACELULAR "EL TERCER SISTEMA"
Description
No tags specified
Slide Set by Víctor Pérez, updated more than 1 year ago
More
Less
|
|
Created by Víctor Pérez
about 4 years ago
|
|
Resource summary
Slide 1
MATRIZ EXTRACELULAR
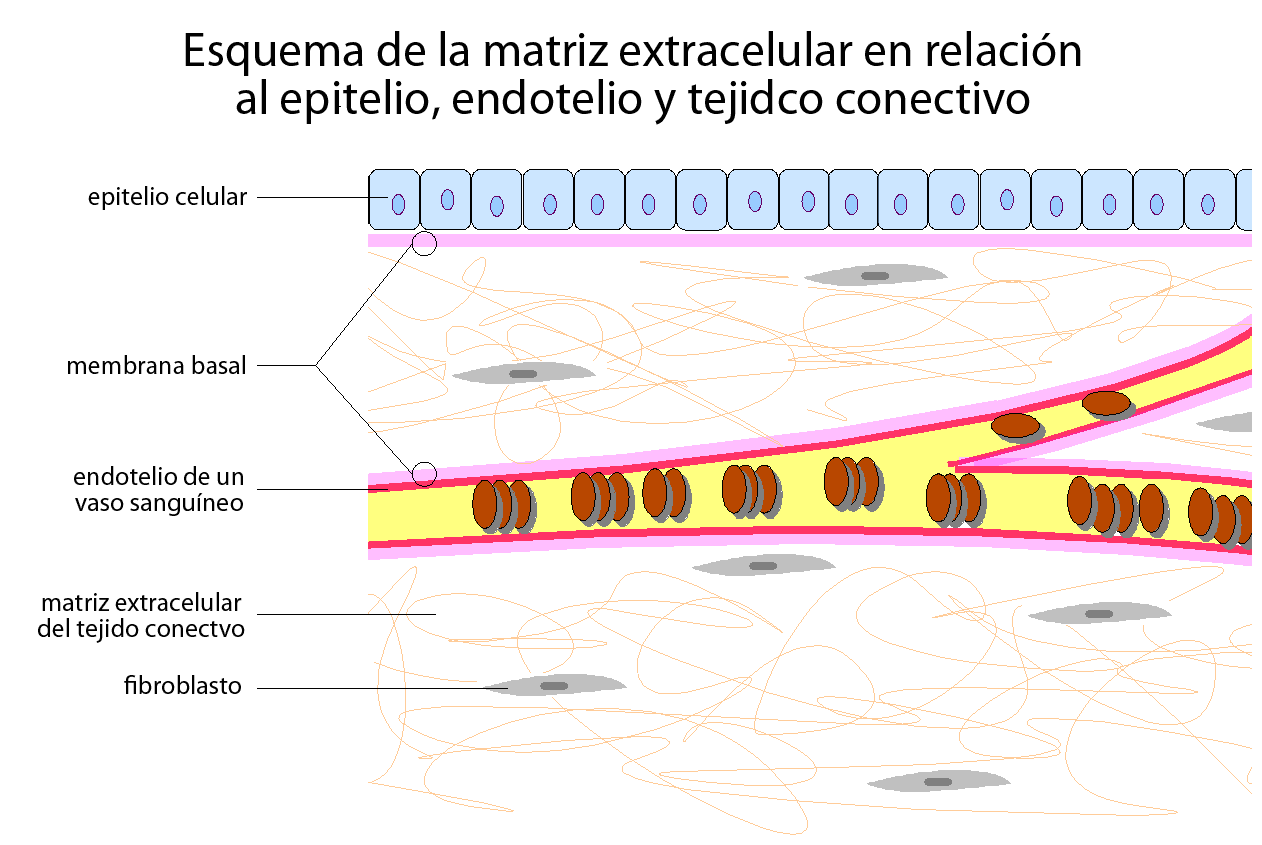
Red tridimensional que engloba todos los órganos, tejidos y células del organismo.
Filtro biofísico de protección, nutrición e inervación celular y el terreno para la respuesta inmune, angiogénesis, fibrosis y regeneración tisular.
Medio de transmisión de fuerzas mecánicas a la membrana basal, que a través de las integrinas soporta el sistema de tensegridad y activa los mecanismos epigenéticos celulares.
El tejido conectivo, una función de comunicación entre órganos, tejidos y células.
Funciones de soporte y comunicación residen en la matriz extracelular (MEC), encargada además del aporte de oxígeno y nutrientes a la célula y eliminación de CO2, toxinas y productos de desecho.
Espacio idóneo para el almacenamiento de material tóxico, especialmente debido a la presencia de proteoglicanos que pueden ligarse fácilmente a una gran variedad de sustancias tóxicas.
En condiciones normales, el acúmulo de toxinas en la MEC es drenado a través del sistema linfático hacia la cadena ganglionar linfática más próxima, y si dicho drenaje no se da o es insuficiente se produce una respuesta que se traduce en inflamación, acidosis y dolor.
Slide 2
Los vertebrados tienen una gran diversidad de tipos de MEC, incluyendo el esqueleto.
La MEC basada en colágeno constituye el principal elemento de soporte estructural de los animales multicelulares, así como desempeña un papel principal en los procesos de organización y orientación tisular, adhesión celular, migración, diferenciación, proliferación y apoptosis.
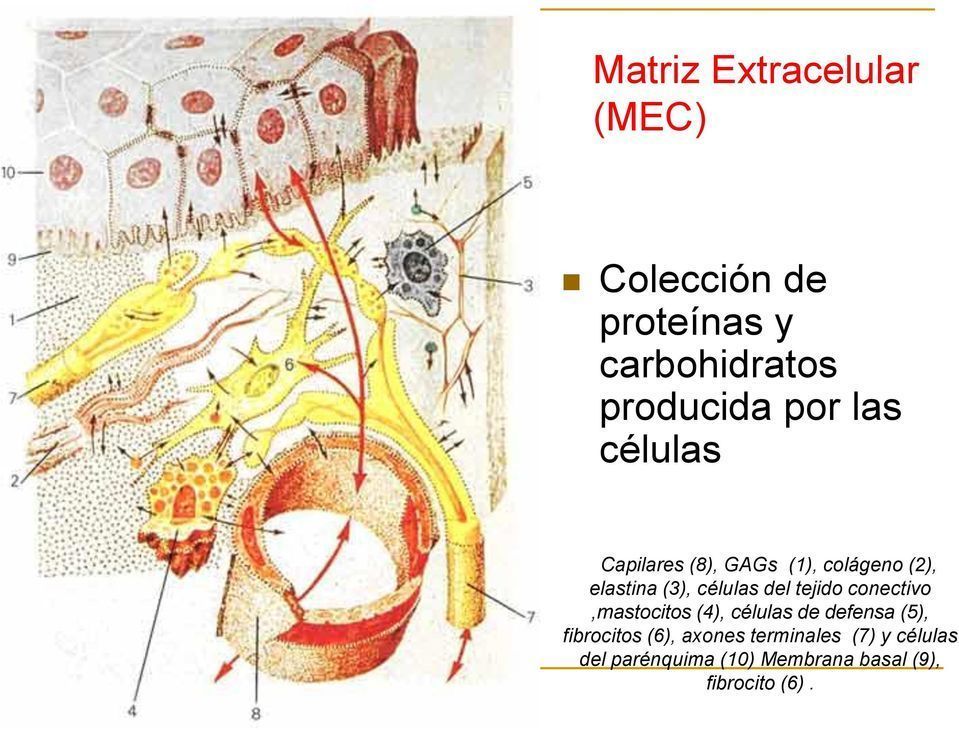
La MEC está compuesta por una compleja mezcla de proteínas, proteoglicanos y glucoproteínas que confieren las propiedades estructurales de células y tejidos.
La MEC activa o deja de hacerlo a los procesos celulares de crecimiento, muerte celular, adhesión, invasión, expresión génica y diferenciación.
La MEC está sujeta a un ritmo dinámico funcional que se activa durante el día y encuentra su fase de reposo en las horas de la noche, dependiente del flujo hormonal que marca el biorritmo de todo el organismo, en especial la hormona tiroidea y el cortisol, aparte del sistema nervioso vegetativo.
{kind=link}
{kind=link}
{kind=link}
Slide 4
El modelo de patología celular de Virchow propone que la unidad mínima de vida en el organismo es la célula. Sin embargo esto es una abstracción, ya que en realidad la célula aislada de su entorno no puede sobrevivir.
Alfred Pischinger expandió este modelo y propuso como unidad mínima de vida en los vertebrados la tríada de capilar-MEC-célula, supone la consideración de la MEC no como un material inerte o un tejido pasivo de sostén, sino como un componente vivo, dinámico y con múltiples funciones, una matriz viviente.
A mitad del siglo XIX, Carl Rokitansky describió el crecimiento del tejido conectivo, afirmando que los elementos celulares y las enfermedades se producían a partir de los líquidos corporales.
Claude Bernard definió el entorno extracelular o el «medio interno del organismo» proponiendo que «su estado saludable era esencial para la salud y que la impureza de este terreno produciría enfermedades»
Slide 5
En el siglo XX, Alfred Pischinger, el padre de la sustancia básica de la MEC, la describe como un gel amorfo que ocupa el entorno extracelular.
Llamó tercer sistema, sistema básico o de regulación basal, a toda la estructura que rodea a la célula, definiéndolo como un sistema de homeostasis.
Los componentes son en general producidos por las mismas células o los aporta la corriente sanguínea. En ambos casos forman el medio donde las células sobreviven, se multiplican y desempeñan sus funciones.
ESTRUCTURA Y FUNCION:
Son de cuatro grandes tipos: (1) sistema colágeno (2) sistema elástico (3) proteoglicanos (4) glicoproteínas multifuncionales (laminina, fibronectina, tenascina, trombospondina y otras).
Los colágenos y el sistema elástico constituyen la arquitectura de la matriz extracelular. Las glicoproteínas actúan como moléculas de adhesión del sustrato intercelular, importantes en las interacciones célula-célula y célula-matriz. Los glicosaminoglicanos y proteoglicanos tienen un papel fundamental en el equilibrio hidroelectrolítico y ácido básico
Slide 6
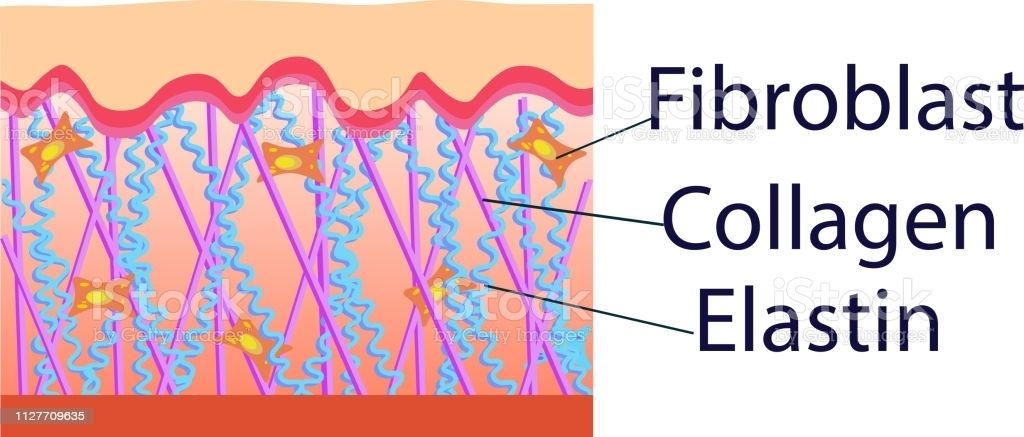
CÓLAGENO
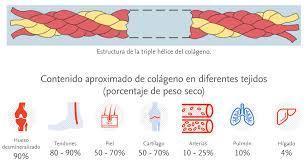
Gran familia de proteínas que tienen por características agruparse formando una estructura supramolecular.
Las moléculas resultan de la asociación de tres cadenas polipeptídicas en con una formación característica de triple hélice.
Poseen dominios globulares, que le confieren flexibilidad y especificidad a las moléculas que los poseen.
Han sido clasificados teniendo en cuenta la forma en que se agregan: colágenos fibrilares I, II, III, V y XI y colágenos no fibrilares VI, VII, VIII, X.
{kind=link}
Slide 7
Los colágenos no fibrilares a su vez se clasifican teniendo en cuenta la constitución y presentación de las fibrillas:
a) Colágenos que forman membrana; son tipo IV, VI y VIII.
b) Colágenos con interrupción de la triple hélice (grupo de FACITS = Fibrila associated collagen whit interrupted triple hélice); son tipo IX, XII, XIV.
c) Colágenos que forman microfibrillas en cuenta de rosario; son tipo VI.
d) Colágeno que forman fibras de anclaje; son tipo VII.
El colágeno tipo I es un heterotrímero formado por dos cadenas α1 (I) y una cadena α2 (I), con 300 nm de longitud. Se encuentra en la dermis, en el tendón, en la lámina propia de todas las mucosas y en la placenta se localiza a nivel del estroma velloso. Es la molécula más abundante, y la encontramos además en los procesos de inflamación crónica y fibrosis, sustituyendo los tejidos parenquimatosos, como fue observado en las vellosidades coriales de la placenta de mujeres con Enfermedad Hipertensiva del Embarazo (EHEE).
El colágeno tipo II es un homotrímero formado por tres cadenas α1 (III) y se encuentra en tejidos que necesitan más elasticidad y menor rigidez, como los tejidos embrionarios y linfáticos. En la vellosidades coriales se localiza debajo del trofoblasto y en el estroma de la placenta
El colágeno tipo III es un homotrímero formado por tres cadenas α-1 (III). Se encuentra en los tejidos que necesitan más elasticidad y menor resistencia como los tejidos embrionarios, linfáticos y formando parte del estroma vellositario normal
Slide 8
El colágeno tipo V es un heterotrímero, presenta tres cadenas α-1 (V), α-2 (V), α3 (V) y se localiza en la piel, el músculo liso y córnea.
El colágeno tipo XI es un heterotrímero formado por las cadenas α-1 (XI), α2 (XI) y α-3 (XI) y se encuentra en el cartílago.
Los colágenos que forman membrana encontramos los tipos IV, VI y VIII.
El colágeno IV se caracteriza por formar una trama filamentosa en forma de una malla conocida como «tela de corral de gallina» y se localiza en las membranas basales. Está formado por dos tipos de cadenas, α-1 (IV) y α-2 (IV), que se asocian para formar una malla tridimensional. En procesos inflamatorios y en cáncer es destruida por la colagenasa producto de los leucocitos y células neoplásicas
El colágeno VI es un heterotrímero formado por tres cadenas genéticamente distintas: α-1 (VI), α-2(VI) y α-3 (VI). Desempeña una función importante en las interacciones entre los diferentes componentes matriciales y en los mecanismos de adhesión y tiene particular importancia en el reconocimiento de la integrina α2β1 que regula la adhesión y diferenciación celular.
Hay colágeno VI en los tendones, piel, cartílago elástico, espacio perisinusoidal del hígado, donde es sintetizado por la células de Ito; , en la cirrosis hepática está aumentado.
Los colágenos tipo IX y XII han sido descritos a nivel del tejido cartilaginoso interactuando con el colágeno II.
El colágeno tipo XIV es un homotrímero con características de FACIT que interactúa con el colágeno I. Se localiza en la piel y tendones.
Slide 9
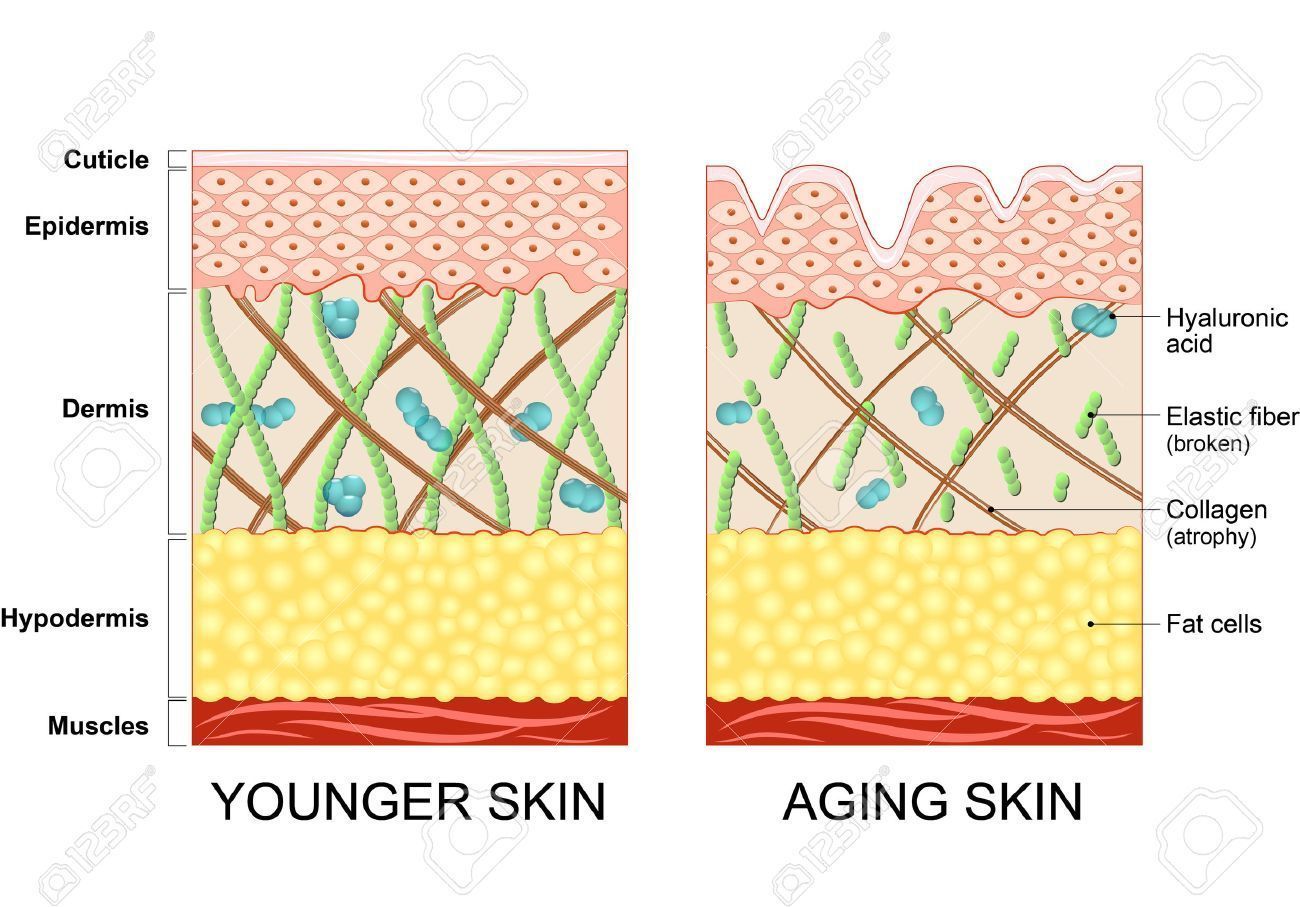
Sistema elástico
Formado por la elastina, que es un polímero insoluble, constituido por moléculas solubles de tropoelastina, y por una glicoproteína denominada fibrilina.
Las microfibrillas se encuentran constituidas por fibrilina. La fibrilina es una glicoproteína formada por un monómero con peso molecular de 350Kd; es rica en cisteína y presenta en su estructura una secuencia semejante al factor de crecimiento epidérmico (EGF) y otra parecida al factor de crecimiento y transformación del fibroblasto (FGβ1)
La fibrilina desempeña una función importante en el alineamiento de las moléculas de tropoelastina, de tal forma que permite la formación de las uniones cruzadas
En las vellosidades coriales, las fibras elásticas se localizan en el estroma velloso y en torno de las células mioepiteliales perivasculares, las cuales regulan el tono y la elasticidad de los vasos coriales, lo cual facilita el flujo sanguíneo entre la madre y el feto.
En la piel, el sistema elástico es fundamental para mantener su elasticidad. La elastina es sensible a la luz ultravioleta y a la exposición al sol, lo cual acelera el proceso de envejecimiento.
Slide 10
{kind=link}
{kind=link}
Slide 11
PROTEOGLICANOS
Los proteoglicanos son complejos de macromoléculas formados por la asociación covalente entre cadenas polipeptídicas y glicosaminoglicanos.
El ácido hialurónico es el único glucosaminoglicano que no se une a la cadena peptídica; tiene un papel muy importante en la migración celular. Facilita la hidratación de los tejidos, debido a la gran cantidad de radicales libres, que se ligan a las moléculas de agua. Por lo tanto, la hidratación de los tejidos depende de la concentración y el estado fisiológico del ácido hialurónico.
el ácido hialurónico al unirse a la proteína B forma el complejo ácido hialurónico proteína B, asociado al estímulo de la actividad de proteína quinasa, que sirve como señal de traducción a nivel celular, y tiene un papel importante en la interacción de la superficie celular con el citoesqueleto.
Función de los proteoglicanos es contribuir a la adhesividad celular mediante su interacción con la superficie celular y con otros componentes matriciales.
El sindecan,, proteoglicano de la membrana celular, transmite señales a proteínas transmembranales, como las integrinas, que a su vez interactúan con el citoesqueleto, el cual facilita la interacción de los filamentos de actina
Slide 12
La unión sindecan a la fibronectina es mediada por el heparan-sulfato
Los proteoglicanos pueden regular la diferenciación y proliferación celular en algunos tejidos, incluidos el cartílago.
El heparan-sulfato perlecan es el primero de los proteoglicanos en aparecer en la lámina basal junto con la laminina y el colágeno IV estableciendo las bases estructurales de las membranas basales
Slide 13
Glicoproteínas multifuncionales
Fibronectina (FN), laminina (LN), tenascina (TN) y trombospondina (TB). Para cumplir con sus funciones estas moléculas necesitan de otras moléculas que sirven de unión entre la matriz extracelular y el citoesqueleto celular, como son: las integrinas, las caderinas, las inmunoglobulinas y las selectinas.
FIBRONECTINA
Glicoproteína de adhesión celular, se encuentra formada por un dímero de unidades idénticas, enlazadas entre sí por puentes disulfuros.
Es una molécula multifuncional que presenta varios dominios globulares, los cuales le dan ciertas características que le permiten ejercer diferentes funciones.
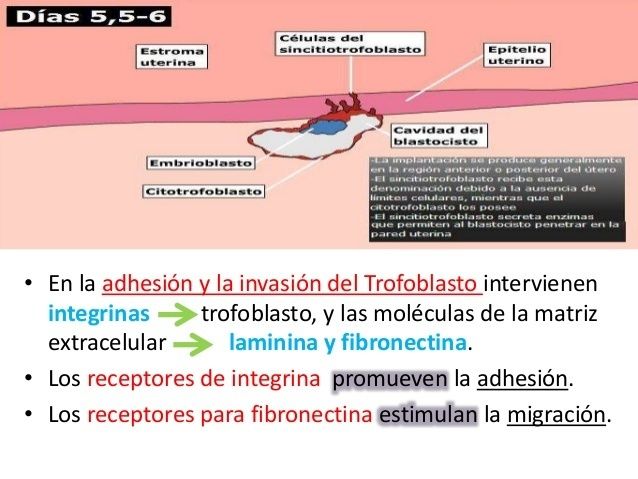
En la placenta, la fibronectina se encuentra en el estroma velloso, en el fibrinoide intra e intervelloso, y parece tener un papel fundamental en la adhesión de las células trofoblásticas a la decidua, se reconocen así los receptores de las integrinas
Slide 14
En el cáncer de cervix, la fibronectina se localiza a nivel de las células neoplásicas y en el estroma, lo que permite a la célula cancerosa no ser reconocida por el sistema inmune.
{kind=link}
{kind=link}
{kind=link}
Slide 16
LAMININA
Glicoproteína que posee un peso molecular de 900.000 kd tiene forma de cruz y está compuesta por tres cadenas polipeptídicas diferentes, denominadas (α1), β1 (β1) y β2(γ1).
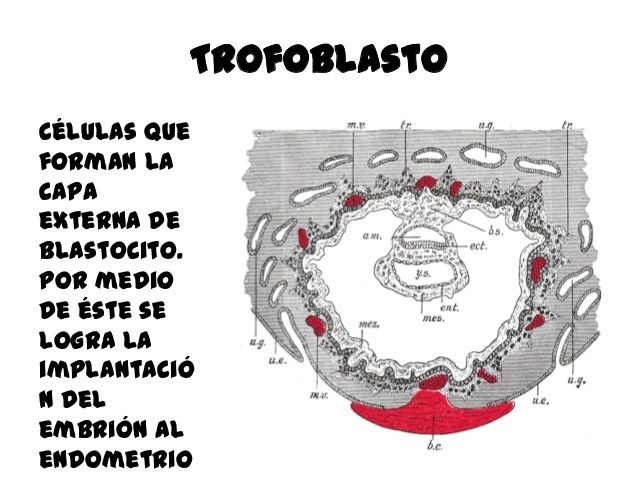
En el trofoblasto, la laminina se localiza en la membrana basal y tiene un papel fundamental durante el proceso de implantación y en la diferencia de las vellosidades placentarias.
MOLECULÑAS DE ADHESION
Grupo de glicoproteínas que tiene como función regular las relaciones entre las células y la matriz extracelular. Son elementos fundamentales en las interacciones célula-célula y célula-matriz.
Presentan peso molecular que varía de 90 a 220 Kd. Son mediadoras de las interacciones célula - célula y célula-matriz
Slide 17
Uniones intercelulares
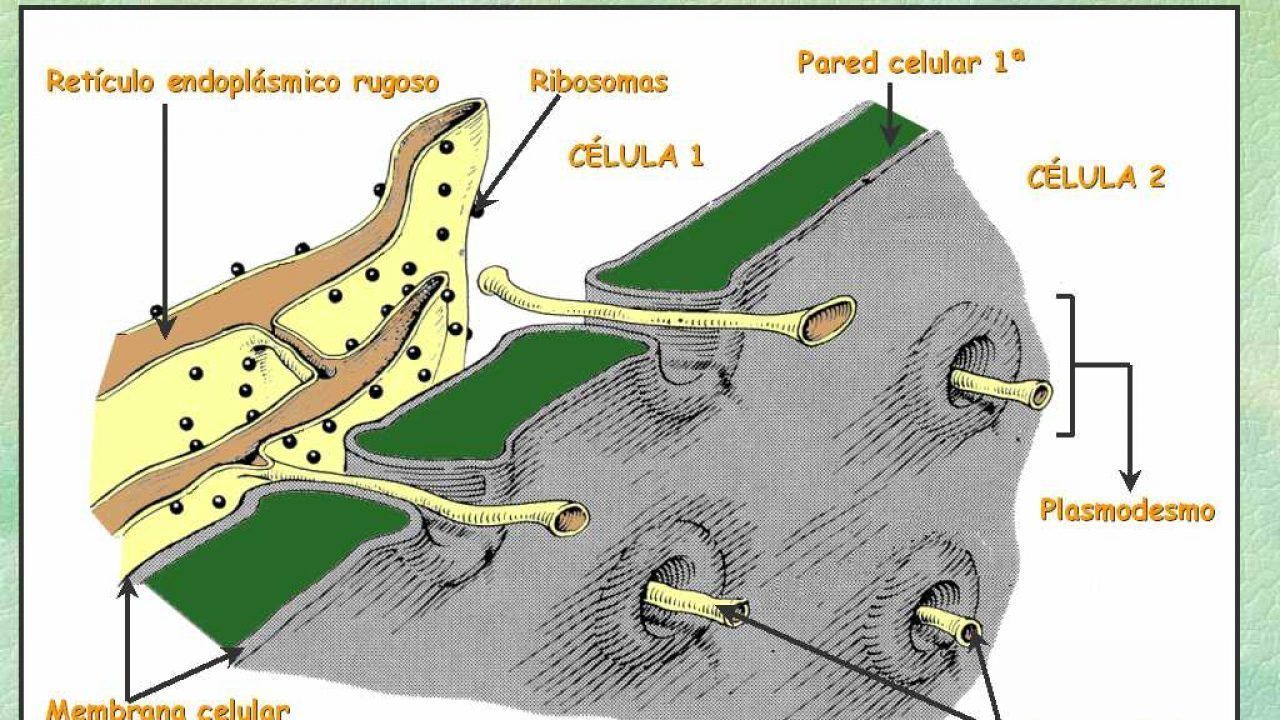
PLASMODEMOS
Las células vegetales, al estar rodeadas de pared celular, no están en contacto unas con otras mediante amplias extensiones de membrana plasmática como las células animales. Sin embargo, poseen uniones especializadas llamadas plasmodesmos (en singular plasmodesmo) en la pared celular que permiten el intercambio citoplásmico directo entre dos células.
Están recubiertos de membrana plasmática que es continua con las membranas de ambas células.
Slide 18
Las moléculas menores a cierto tamaño (conocido como límite de exclusión por tamaño) se mueven libremente a través del canal del plasmodesmo por medio de difusión pasiva.
Los plasmodesmos pueden dilatarse (expandirse) de manera selectiva para permitir el paso de ciertas moléculas grandes, como proteínas, aunque este proceso no está bien comprendido^{1,2}
{kind=link}
Slide 19
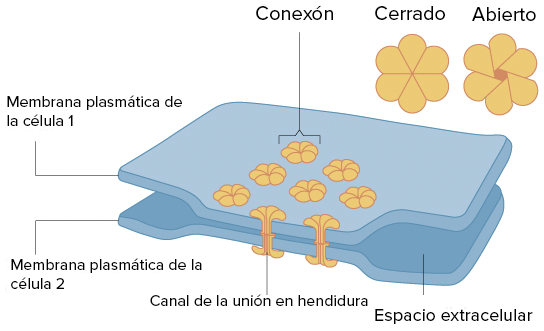
UNIONES EN HENDIDURA
Son canales entre células vecinas que permiten el transporte de iones, agua y otras sustancias.
En los vertebrados, la uniones en hendidura se forman cuando un conjunto de seis proteínas de la membrana, llamadas conexinas, forman una estructura alargada parecida a una dona llamada conexón.
Los invertebrados forman uniones en hendidura similares pero usan un conjunto de proteínas diferentes llamadas inexinas.
Importantes en el músculo cardíaco: la señal eléctrica que induce la contracción se propaga rápidamente entre las células del corazón a medida que los iones pasan a través de las uniones en hendidura, lo que permite que las células se contraigan de manera simultánea
{kind=link}
Slide 20
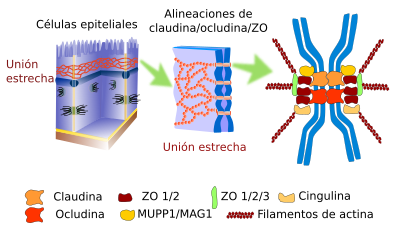
UNIONES ESTRECHAS
Las uniones estrechas crean un sello a prueba de agua entre dos células animales adyacentes.
las células se mantienen unidas firmemente por muchos grupos individuales de proteínas de unión estrecha conocidas como claudinas.
El propósito de las uniones estrechas es evitar que el agua escape entre las células, lo que permite que una capa de células (como las que recubren un órgano) actúe como una barrera impermeable. Por ejemplo, las uniones estrechas entre las células epiteliales que recubren la vejiga evitan que la orina escape hacia el espacio extracelular.
{kind=link}
Slide 21
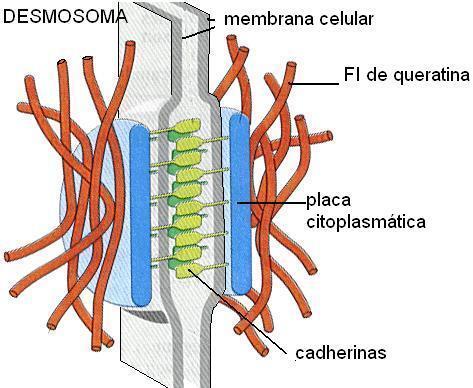
DESMOSOMAS
Actúan como puntos de soldadura entre células epiteliales adyacentes. Un desmosoma está compuesto de un complejo de proteínas, algunas de las cuales se extienden a través de la membrana, mientras que otras anclan la unión dentro de la célula.
Las cadherinas, proteínas de adhesión especializadas, se encuentran en las membranas de ambas células e interactúan en el espacio entre ellas, y las mantiene unidas.
{kind=link}
Want to create your own Slides for free with GoConqr? Learn more.