18440904
Descripción
Fichas por Nele Tatarevic, actualizado hace más de 1 año
|
|
Creado por Nele Tatarevic
hace más de 5 años
|
|
| Pregunta | Respuesta |
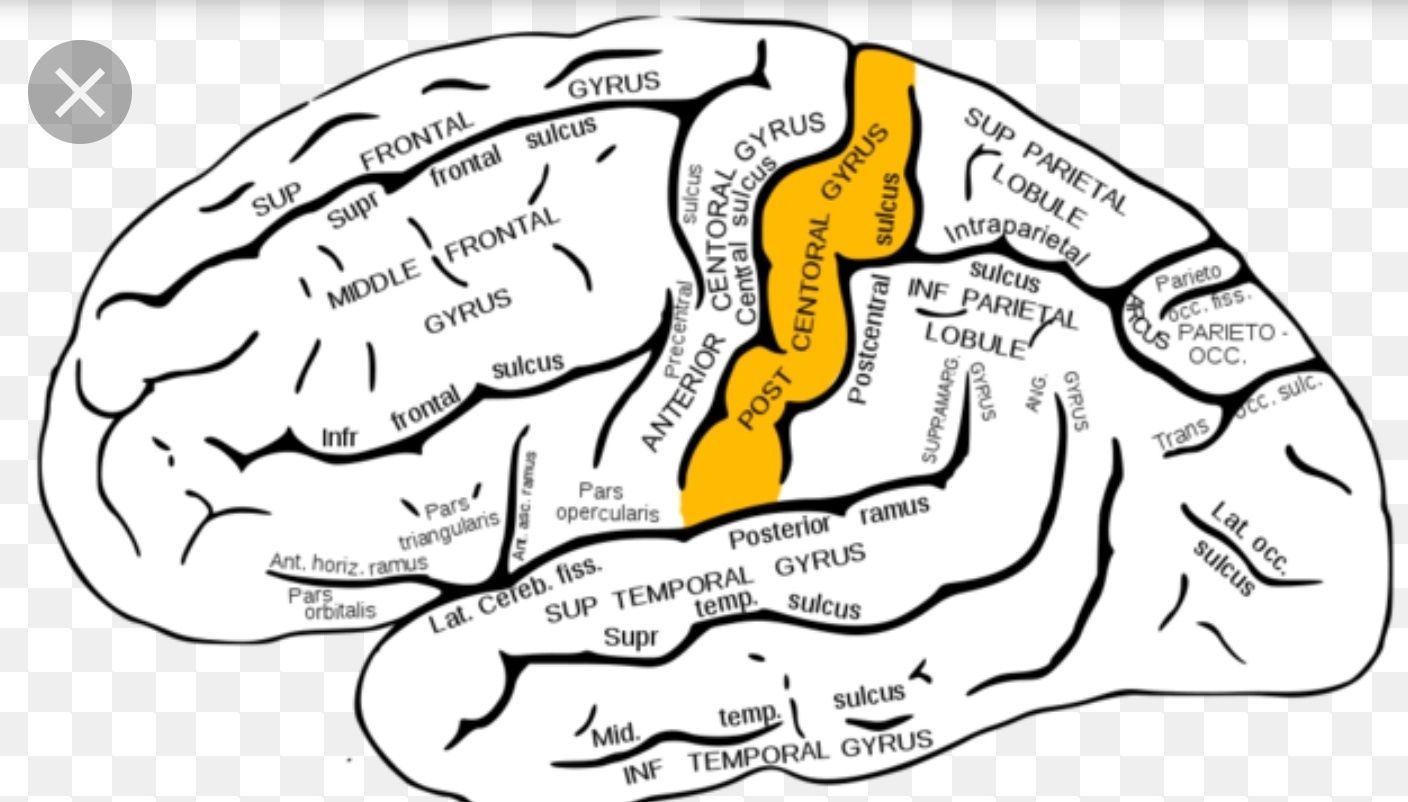
| Kapitel 5: Erläutern Sie die Anordnung des somatosensorischen Homunculus. | - liegt auf dem Gyrus Postcentralis (primärer somatosensorischer Cortex) - somatotop angeordnet = nebeneinander liegende Punkte auf der Haut sind auch nebeneinander auf dem Cortex abgebildet - von medial nach lateral: Zehen, Fuß, Beine, Hüfte, Genitalien, Rumpf, Kopf (ohne Gesicht), Schultern, Arme, Hand, Finger, Zunge, Gesicht und Eingeweide --> Hände, Zunge, Genitalien und Lippen sind vergrößert —> weil dort die taktilen Rezeptoren besonders dicht verteilt sind —> in diesen Bereichen kann man Berührungen besonders gut orten &kleinste Eigenschaften eines uns berührenden Objekts erkennen |
| Kapitel 5: Wie konvergiert unser Gehirn Gerüche in ein Aktivierungsmuster? | 1.) Gerüche gelangen in die Nase 2.) Geruchsstoff trifft auf olfaktorische Zellen der Riechschliemhaut 3.) olfaktorische Zellen werden mithilfe von Zilien durch Duftstoffe aktiviert 4.) Zilien haben Geruchsrezeptoren (ca. 350 verschiedene) 5.) Die Signale der Axone werden durch das Siebbein zum Bulbus Olfactorius geleitet 6.) dort bilden die Axone Synapsen mit den Glomeruli 7.) Glomeruli sind kugelförmige Anordnungen von Neuronen, die nur Input von Geruchszellen, die den gleichen Rezeptor enthalten 8.) Die Info eines Glomerulus wird dann über Mitralzellen zum Gehirn zu Geruchsregionen weitergeleitet 9.) eine gleichzeitige Aktivierung verschiedener Glomeruli erzeugt ein Muster, welches vom Bulbus Olfactorius dekodiert wird & als bestimmter Geruch identifiziert wird (z.B. Kaffee) 10.) hier wird also ein nicht-topografisches Signal in ein topografisches Signal übersetzt --> es entsteht eine sensorische Landkarte für Gerüche im Gehirn 11.) wird die Info an den Cortex piriformis weitergeleitet , sind die Landkarten vermisch |
| Kapitel 5: Welche Cortexarealtypen gibt es? | 1.) Primär sensorische Areale 2.) Assoziativ-sensorische Areale 3.) Multimodale Areale 4.) Prämotorische Areale 5.) Primär motorische Areale |
| Kapitel 5: Wie funktionieren primär sensorische Areale? | - sensorische Information wird sehr wenig aufgearbeitet repräsentiert - im V1 sind das z.B. Visuelle Punkte, Kanten, Signale einer bestimmten Wellenlänge usw. - visueller Pfad teilt sich in 2 Richtungen: a) dorsaler Pfad: erreicht Parietalcortex —> wichtig für räumlich-visuelle Verarbeitung & Planung visuomotorischer Prozesse (visuell-räumlich) b) ventraler Pfad: erreicht Temporalcortex —> für komplexere Analyse visueller Objekte zuständig (visuell-objektbasiert) - Verletzungen dieser Areale führen zu Defiziten in der bewussten Wahrnehmung dieses Sinnes --> Bsp.: Blindsehen --> Bsp.: bei Läsion des primären auditorischen Cortex sind Patienten weitgehend gehörlos --> Bsp.: bei Läsion des somatosensorischen Cortex: Patienten können die Rauhigkeit von Gegenständen nicht mehr mit ihrer contralateral zur Läsion liegenden Hand ertasten können |
| Kapitel 5: Wie funktionieren assoziativ-sensorische Areale? | - hier projizieren Neurone aus den primär sensorischen Arealen hin - Information wird hier mehr und mehr zu haptischen, auditorischen oder visuellen Objekten verknüpft - aus unverarbeiteten sensorischen Elementen entstehen echte Objektrepräsentation - Bei Läsion des somatosensorischen Assoziationscortex: Patienten können einfache sensorische Leistungen (z.B. Farberkennung, Geräuschlokalisation) erbringen, aber Objekte im ganzen nicht erkennen - Bei Läsion in assoziativ-visuellen Arealen im Temporalcortex: visuelle Agnosie |
| Kapitel 5: Wie funktionieren multimodale Areale? | - entstehen durch das Zusammenschalten der Informationsströme verschiedener Modalitäten - diese mulitmodalen Repräsentationen werden in den Assoziationsarealen zusätzlich mit motivationalen und emotionalen Infotmationen assoziiert - die größten multimodalen Areale liegen im: Temporal-, Parietal- und Frontalcortex --> visuelle Signale erreichen alle 3 Assoziationsareale --> Im Temporalcortex werden die visuellen Infos mit auditorischen Signalen verknüpft und schaffen Grundlage für unser Sprachsystem --> Assoziationsregion im Parietalcortex verarbeitet die Lokalisation von Gegenständen im Raum sowie die Berechnung von Eigenbewegungen im dreidimensionalen Raum. Hier entstehen auch arithmetische Denkprozesse, z.B. Addieren --> wir repräsentieren Zahlen als eine mentale Reihe, also als Zahlenstrahl. Wir transformieren numerische Werte in räumliche Positionen. --> dort entstehen Wort- und Raumverarbeitungen und Handlungsplanungen --> größtes Assoziationsareal ist der PFC. Weitere infos Kapitel 6 - bei Läsion: Defizite in den jeweiligen kognitiven Leistungen |
| Kapitel 5: Wie funktionieren prämotorische Areale? | - Assoziationsareale können bei Handlungsintention Zellverbände aus prämotorischen Arealen aktivieren - diese Zellverbände sind spezialisiert auf bestimmte Bewegungstypen und - richtungen - Zellen starten Bewegung nicht, sondern wählen sie aus - prämotorische Areale wählen Handlungsoptionen aus anhand von externen Signalen, wie z.B. ein Ton - bei Läsion: Auf Kommando können Hanldungen nicht mehr ausgeführt werden, spontan aber schon - der Prämotorcortex auf der Innenseite des Cortex trifft die Handlungsauswahl entsprechend der Intentionen der Person und nicht entsprechend externer Signale |
| Kapitel 5: Wie funktionieren primär motorische Areale? | - prämotorische & multimodale Areale projizieren zum primären motorischen Cortex (=letzte corticale Station beim Ausführen einer Handlung) - von hier führt die Pyramidalbahn in das Rückenmark und in die motorischen Hirnnervenkerne, um Handlungen zu initiieren - auf dem Motorcortex gibt es einen auf dem Kopf stehenden motorischen Homunculus, der die Muskelgruppen entsprechend ihrer Nachbarschaft repräsentiert --> Hand und Mund sind stark vergrößert - bei Läsion: Lähmungen in contralateralen Extremitäten, wobei rumpfferne Muskeln immer stärker betroffen sind als rumpfnahe |
| Kapitel 5: apperzeptive Agnosie | - bei Verletzung im posterioren Bereich des visuellen Assoziationsareals des Temporalcortex - der ventrale Pfad, in dem visuelle Objektverarbeitungsprozesse stattfinden, ist partiell gestört, sodass ein Abschluss des Perzeptionsprozesses nicht möglich ist - einzelne Aspekte eines Gegenstandes können erkannt werden, aber nicht das ganze - Patienten erkennen keine Gegenstände und können sie nicht abzeichnen, wohl aber Gegenstände aus dem Gedächtnis zeichnen - der Perzeptionsprozess ist noch nicht abgeschlossen - sie können keine Formen diskriminieren |
| Kapitel 5: Assoziative Agnosie | - Läsion liegt im vorderen Bereich des visuellen Assoziationsareals des Temporalcortex - Gesamtheit eines Gegenstandes kann nicht erkannt werden, aber Einzelheiten - hier ist Perzeptionsprozess abgeschlossen - sie können Gegenstände abzeichnen - sie können keine Gegenstände aus dem Gedächtnis abzeichnen - visuelle Vorstellung ist gestört - sie können Formen diskriminieren - entstehen wahrscheinlich durch eine Verletzung am Endpunkt der ventralen Bahn --> dadurch können die fertigen Bilder nicht mehr mit dem Weltwissen des Restgehirns kombiniert werden - Patienten erkennen Objekte aber, wenn sie andere Sinne anwenden, z.B. Fühlen - Patienten erkennen Objekte, indem sie jede relevante Information nutzen, die sie aus dem Bild ziehen können (z.B. ringeliger Schwanz --> es muss ein Schwein sein) --> deshalb erkennen sie Dinge am besten, wenn das Bild besonders viele Informationen liefert --> reale Objekte, dann Fotos, dann Zeichnungen |
| Kapitel 5: Blindsehen bei unilateraler Läsion des primären visuellen Cortex | Wenn man die Patienten darum bittet, in ihrem contralateralen blinden visuellen Halbfeld zu greifen, können sie trotz bewusster Blindheit sehr gut danach greifen |
| Wie funktioniert der Pfad für visuelle Leistungen und bewusstes Sehen? ... und der Pfad, der evtl für Blindsehen verantwortlich ist | - Retina - Corpus geniculatum laterale pars dorsalis - primärer visueller Cortex (Brodmann-Area 17) - entlang des ventralen Pfades den Temporalcortex entlang noch ein Pfad: - Retina - Superior colliculus des Mittelhirns - thalamisches Pulvinar - visuelle corticale Areale außerhalb der Area 17 - diese Route dominiert vor allem die Projektion entlang des dorsalen Pfades zum Parietalcortex --> eventuell ist dieser Pfad für Blindsehen verantwortlich |
| Wie viele Geruchsrezeptoren besitzen olfaktorische Zellen? | Ca. 350 Das heißt, so viele unterschiedliche primäre Geruchsstoffe können wir wahrnehmen |
| Untersuchung von Isaacs et al. (2001) zu Dyskalkulie | So untersuchten z. B. Isaacs et al. (2001) die Gehirne von Erwachsenen, die zu früh geboren wurden und von denen die Hälfte unter Dyskalkulie (Rechenschwäche) litt. Diejenigen Personen mit Rechenproblemen hatten eine Reduktion ihrer grauen Substanz in einem Bereich des parietalen Assoziationscortex der linken Hemisphäre. --> die ist wichtig für arithmetische Prozesse |
| Experiment zu prämotorischen Arealen an Rhesus-Affen | Experimenten, bei denen Rhesusaffen trainiert werden, bei einem Tonsignal einen Hebel zu bewegen. Einige Sekunden vor dem Ton wird den Tieren durch ein Farbsignal angezeigt, ob gleich der Hebel gezogen oder gedrückt werden muss. Die Zellen des Prämotorcortex werden in einer solchen Aufgabe durch das Farbsignal aktiviert (Moment der Handlungsauswahl) und nicht durch den Ton (Beginn der Bewegung). Der Prämotorcortex auf der Lateralseite des Cortex wählt Handlungen entsprechend externer Signale aus. |
| Kapitel 5: Wieso konnte Frau Buchner sich problemlos zwischen Möbeln bewegen ohne sie anzustoßen? | Bei Frau Buchner ist der primäre visuelle Cortex intakt (primäre visuelle Eindrücke werden verarbeitet) und ihr dorsaler visueller Pfad in den Parietalcortex ist weitestgehend funktionsfähig (räumliche Orientierung gelingt). |
| Kapitel 5: Was ist Blindsehen und wie testet man es? | noch mit Buch und Vorlesung abgleichen Blindsehen bezeichnet die Fähigkeit, Objekte zu lokalisieren und sie zu begreifen, obwohl die betroffene Person keine bewusste Repräsentation des Gesehenen besitzt und sich als blind empfindet. Die Läsion umfasst i. d. R. den okzipitalen Cortex mit dem Brodmann-Areal 17. Wahrscheinlich wird das Blindsehen durch die retinale Projektion über den Superior colliculus des Mittelhirns zum thalamischen Pulvinar und von dort zu visuellen corticalen Arealen außerhalb des Areals 17 geleistet. Man kann es testen, indem man Patienten bittet, nach einem Gegenstand zu greifen oder einen Laserpointer mit den Augen zu verfolgen. Die Patienten werden diese Aufgaben adäquat lösen. |
| Kapitel 5: Wie erfolgt eine Aufmerksamkeitsverschiebung auf thalamischer Ebene? | noch mit Buch und Vorlesung abgleichen Ausgangszustand: Die Zelle im Tegmentum ist Teil des pontomesencephalotegmentalen Komplexes (PMT). Sie ist acetylcholinerg und projiziert in den somatosensorischen Thalamus. Das Neuron im Thalamus repräsentiert den rechten Fuß. Es wird von einem GABAergen Interneuron gehemmt und feuert dementsprechend sehr wenig. Dadurch überträgt es nur kaum Aktivität auf das kortikale Pyramidalneuron, welches zusätzlich durch ein weiteres Interneuron gehemmt wird. Das widerum führt zur geringen Aktivität des PMT. Man spürt den Fuß nicht. Nach Erwähnung: Das Interneuron im Kortex, welches zuvor des Pyramidalneuron gehemmt hat, wird nun durch Neurone aus präfrontalen Arealen gehemmt. Damit ist das Pyramidalneuron aktiv und kann die Aktivität über glutamaterge Rezeptoren auf die Zelle im PMT übertagen. Dieses kann nun die Interneurone hemmen, sowie die Zelle im Thalamus aktivieren. Somit spürt man den Fuß. |
| Kapitel 5: Was ist ein Homunculus? | Bezeichnete ursprünglich einen künstlich geschaffenen Menschen. Heute bezeichnet ein Homunculus in der Neurowissenschaft die Repräsentation des Körpers auf dem somatosensorischen bzw. motorischen Cortex. Die Verzerrung der Repräsentation ensteht durch die ungleiche Verteilung der taktilen Rezeptoren auf unserer Haut. Damit sind diese auch in unserem Cortex unterschiedlich stark repräsentiert, sodass die Hände sowie Lippen und Zunge grotesk vergrößert sind. - konnte kartiert werden duch Wilder Penfield durch Mikrostimulation am Gehirn |
| Kapitel 5: Wie erfolgt eine Aufmerksamkeitsverschiebung auf thalamischer Ebene? | Ausgangszustand: Die Zelle im Tegmentum ist Teil des pontomesencephalotegmentalen Komplexes (PMT). Sie ist acetylcholinerg und projiziert in den somatosensorischen Thalamus. Das Neuron im Thalamus repräsentiert den rechten Fuß. Es wird von einem GABAergen Interneuron gehemmt und feuert dementsprechend sehr wenig. Dadurch überträgt es nur kaum Aktivität auf das kortikale Pyramidalneuron (was für den Fuß zuständig ist), welches zusätzlich durch ein weiteres Interneuron gehemmt wird. Das wiederum führt zur geringen Aktivität des PMT und so kann nur wenig Aktivität in den Thalamus übertragen werden. Man spürt den Fuß nicht. Nach Erwähnung: Das Interneuron im Kortex, welches zuvor des Pyramidalneuron gehemmt hat, wird nun durch Neurone aus präfrontalen Arealen gehemmt. Damit ist das Pyramidalneuron aktiv und kann die Aktivität über glutamaterge Rezeptoren auf die Zelle im PMT übertagen. Dieses kann nun die Interneurone hemmen, sowie die Zelle im Thalamus aktivieren. Somit spürt man den Fuß. |
| Kapitel 6: Erläutern Sie die funktionelle Organisation des präfrontalen Cortex. | - größter Teil des Frontalcortex - steuert alle höheren exekutiven Funktionen - der laterale PFC lässt sich entlang der anteroposterioren und der dorsoventralen Achse in verschiedene funktionelle Zonen einteilen posteriore Achse (von posterior nach anterior): - regelbasierte Wahl von Handlungsalternativen (Areal 8) - Übrewachung zeitgleich ablaufender Prozesse (Areale 9 und 46) - Kontrolle der Kontrolleure (Areal 10) dorsoventrale Achse: - Kontrolle kompllexer Ablaufprozesse (Areale 44, 45 und 47) - Entscheidungen entsprechend dem subjektiven Wert der Alternativen (Areale 11-14 und 25) - Evaluation eigener Leistungen und soziale Kognition (Areal 24 und 32) --> zu den einzelnen Arealen noch Karteikarten machen? |
| Kapitel 6: Wie werden Handlungen von den Arealen 8 und 6 organisiert? | Das Brodmann-Areal 8 gehört dem prämotorischen Cortex an, Areal 6 hingegen nicht, und sie beteiligen sich an der Handlungsauswahl. Patienten mit Läsionen in diesen Regionen haben keine Probleme, eine Handlung motorisch auszuführen. Sie haben auch kein Problem, die Instruktion für eine Aufgabe kognitiv zu verstehen. Trotzdem gelingt es ihnen nicht, auf den jeweiligen Stimulus und entsprechend der Instruktion die richtige Handlung auszuwählen. Kein Problem verursachen dagegen konditionierte Reaktionen auf hochgradig überlernten Stimulus-Reaktions-Assoziationen. |
| Kaptel 6: Was ist ein Ensemble? | Gruppen von Neuronen, die durch ihre gemeinsame Aktivität ein Objekt oder einen Gedanken repräsentieren. Ensembles sind somit temporäre Koalitionen von aktiven Neuronen. Diese Koalitionen entstehen, zerfallen und bilden sich in veränderter Zusammensetzung aufs Neue; genau wie die Gedanken in unserem Kopf. Sie können voneinander unabhängig sein, obwohl einzelne Neurone Mitglieder mehrerer Ensembles sind. - haben starke synaptische Verbindungen untereinander - Neurone eines Ensembles haben stärkere synaptische Verbindungen zu Neuronen innerhalb ihres Ensembles als zu solchen anderer Ensembles |
| Kapitel 6: Kann man im EEG lokale und weit verstreute Ensembles unterscheiden? | -Wenn sich ein lokales Ensemble bildet, feuern alle Neuronen dieser Gruppe und aktivieren sich ununterbrochen gegenseitig. Wenn im EEG hochfrequente örtlich begrenzte Aktivität zu sehen ist, kann das ein Zeichen für ein lokales Ensemble sein. Die Streuung muss aber groß genug sein, um die verschiedenen lokalen Maxima mit der eher schlechten räumlichen Auflösung des EEG differenzieren zu können. - das Wechseln der Zugehörigkeit individueller Neurone zwischen verschiedenen Ensembles abhängig von kognitiven Prozessen lässt sich im EEG nicht erkennen weiter im Buch lesen auf S. 195 |
| Kapitel 6: Wie ist die Wirkung der Aktivierung dopaminerger Rezeptoren auf das Wachsen und Zerfallen von neuronalen Ensembles? | Noch mit Buch und Vorlesung abgleichen Insgesamt gibt es fünf verschiedene Dopaminrezeptoren, die in D1- und D2-Rezeptoren aufgeteilt werden. Eine massive Ausschüttung von Dopamin aktiviert die D1-Rezeptoren von postsynaptischen Neuronen. Wenn diese Neurone gerade in einem Ensemble aktiv sind, haben sie in diesem Moment sowieso gerade ein hohes Aktivitätsniveau. Die Bindung von Dopamin an D1-Rezeptoren erhöht das neuronale Aktivitätsniveau des Ensembles noch weiter. Benachbarte Nervenzellen, die in diesem Moment keine wichtige Information kodieren, sind dagegen nur wenig aktiv. Die Aktivierung der D1-Rezeptoren hemmt sogar diese Neuronen. Als Konsequenz entsteht wahrscheinlich ein hoher Signal-Rausch-Abstand für den Gedanken, der im Ensemble aufrechterhalten wird. Werden dagegen die D2-Rezeptoren aktiviert, z.B. durch Absinken des Dopaminspiegels im PFC (D2-Rezeptoren können auch bei niedriger Dopaminkonzentration aktiviert werden), werden eher die niedrig aktiven Neurone erregt. Dadurch brechen Ensembles auseinander und neue Gruppen gemeinsam aktiver Neurone können entstehen. |
| Kapitel 6: Wie justiert die dopaminerge Rückmeldung die Synapsen von Neuronen, die an einer Handlung beteiligt sind? | Die Menge und der Zeitpunkt der Freisetzung von Dopamin im Striatum meldet die Güte unserer Vorhersage für das Ergebnis unserer Handlung. Daher werden Lernprozesse über Erwartungsrückmeldungen organisiert. Hierbei wird die Dopaminfreisetzung reduziert, wenn das Ergebnis der Handlung schlechter war, als man erwartet hatte. Ist man dagegen positiv überrascht worden, erhöht sich augenblicklich die Dopaminfreisetzung. Diese sofortige negative oder positive dopaminerge Rückmeldung schwächt die gerade aktiven Synapsen bei Misserfolg oder stärkt sie bei Erfolg. Die neuronalen Verbindungen werden solange modifiziert, bis eine Handlung optimal läuft. Aus diesen Erkenntnissen wurde die Temporal-Difference-Learning-Theorie entwickelt, die einen Mechanismus beschreibt, wie neuronale Rückmeldungen immer bessere Prädiktionen der eigenen Handlungen ermöglichen Somit entsteht im Kreislauf zwischen Cortex und Basalganglien das Gedächtnis für eine Handlung, die wir von nun an in automatisierter Form abrufen können. |
| Kapitel 6: Was ist ein ereigniskorreliertes Potenzial? | Ein ereigniskorreliertes Potenzial (EKP) entsteht aus der Aufmittelung des EEG-Signals nach einem immer wiederkehrenden Ereignis. Dieses Ereignis kann ein Stimulus sein, den man der Versuchsperson immer wieder gibt. Das wiederholte Ereignis kann aber ein mentales Ereignis sein, welches regelmäßig und zeitlich präzise bestimmbar auftritt. Das „Ne“ (Error Negativity) ist ein EKP, das nach dem Auftreten einer fehlerhaften Reaktion auftritt und in einer Interaktion von Basalganglien und medialem PFC generiert wird (fronto-striatales Netzwerk).Die „Ne“ ist also wichtig für die Evaluation eigener Handlungen. Die „Ne verändert sich auch bei neurodegenerativen Prozessen (z.B. Alzheimer). Neurale Systeme verändern sich durch lernabhängige synpatische Plastizität und die Geschwindigkeit dieser Variationen hängt von der individuellen genetischen Ausstattung ab. Deshalb kann man viele Fragestellungen zu neurogenetischen Mechanismen kognitiver Prozesse durch die „Ne“ erforschen. |
| Kapitel 6: Welche Frequenzbänder werden im EEG definiert und mit welchen mentalen Vorgängen werden Sie assoziiert? | Alpha-Band (8-12Hz): - wenn man Augen schließt und müde ist Beta (14-25 Hz): konzentrierte mentale Arbeit Theta (4 bis 7 Hz): Übergang zwischen Wachsein und Schlaf. Bei Tierexperimenten auch bei Aktivität des Hippocampus bei räumlicher Orientierung Delta (<3 Hz): Tiefschlaf Gamma (>25 Hz): könnte Aktivierung eines Ensembles aufzeigen |
| Kapitel 7: Welche Gedächtnistypen gibt es? | - Kurzzeitspeicher - Langzeitspeicher - Arbeitsgedächtnis - Episodisches Gedächtnis - Semantisches Gedächtnis - Prozedurales Gedächtnis |
| Kapitel 7: Welche Eigenschaften hat der Kurzzeitspeicher? | Wird durch Veränderungen der Aktivitäten von Ensembles von Neuronen realisiert; sofort verfügbar, aber kurzlebig passiver Speicher (im Gegensatz zum Arbeitsgedächtnis) --> hier wird Information einfach nur vorgehalten, ohne dass etwas mit dieser Information passiert Das KZG (und das Arbeitsgedächtnis) besteht aus 2 Neuronengruppen: 1.) Eine aktive Gruppe, die den Stimulus repräsentiert. --> sensorische Assoziationscortices 2.) präfrontale Neuronengruppe, die die Neuronen im sensorischen Assoziationscortex organisiert und aktiv hält (ihren Zerfall verhindert) --> Teil der präfrontalen Neurone feuert, wenn der Reiz, der zu merken ist, abwesend ist |
| Kapitel 7: Welche Eigenschaften hat das Arbeitsgedächtnis? | Ein neurokognitives System, mit dem wir eine kleine Menge an Informationen für einen kurzen Zeitraum speichern und bearbeiten können. Wird durch ein aktives Ensemble präfrontaler Neurone realisiert, die Aufmerksamkeit auf diejenigen corticalen Zellen halten, die den relevanten Stimulus verarbeiten Der Hippocampus überführt Arbeitsgedächtnisinhalte in das LZG. |
| Kapitel 7: Welche Eigenschaften hat das episodische Gedächtnis? | Erinnerungen an biografische Ereignisse, die sich so nur einmal abgespielt haben --> episodisches Gedächtnis ist singulär; Der Speicherort ist der Cortex, für Bildung und Abruf des episodischen Gedächtnisses ist der Hippocampus verantwortlich. . Sowohl das episodische als auch das semantische Gedächtnis können leicht verbalisiert werden (explizites, deklaratives Gedächtnis). Um LZG zu bilden, muss das Gehirn morphologisch die Synapsen von Zellverbindungen umbauen |
| Kapitel 7: Welche Eigenschaften hat das semantische Gedächtnis? | Langfristig gespeichertes Faktenwissen. Z.B. Rom ist die Hauptstadt Italiens. Primärer Speicherort ist der Cortex. Zum Aufbau dieses Gedächtnisses ist der Hippocampus relevant. Zum Abruf ist der Hippocampus wahrscheinlich weniger bedeutsam |
| Kapitel 7: Welche Eigenschaften hat das Prozedurale Gedächtnis? | Es speichert unsere motorischen und gedanklichen Fertigkeiten und Routinen (die über Reiz-Reaktions-Ketten gelernt wurden), entsteht durch Übung und ist hauptsächlich in den Synapsen unserer Basalganglien gespeichert. Es entsteht durch Interaktionvon Cortex und Basalganglien. Der Hippocampus ist egal für prozedurales Gedächtnis. Prozedurale Gedächtnisinhalte können ohne Beteiligung des Bewusstseins genutzt werden. --> Bsp.: Schwimmen, flüssig Sprechen Nicht oder nur sehr schwer verbalisierbares (nicht deklarativ, implizit) Gedächtnis. 3 Schritte bis zur Konsolidierung: 1.) kognitive Stufe: konzentriert, latentes Gefühl, man könnte etwas falsch machen, man muss sich verbal an das zu Lernende erinnern 2.) assoziative Stufe: man kennt die Schritte nun, trotzdem Konzentration nötig 3.) Autonome Phase: man kann der Handlung nachgehen, während man andere Dinge macht In der 2. Nachthälfze wird prozedurales Gedächtnis konsolidiert. |
| Ordne die Begriffe deklarativ, nicht deklarativ, verbalisierbar, explizit und implizit | deklarativ = verbalisierbar = explizit nicht deklarativ = nicht verbalisierbar = implizit |
| Kapitel 7: Was passiert während der Aktivierung des Arbeitsgedächtnisses im präfrontalen Cortex? | noch nicht abgeglichen Ergebnisse aus Einzelzellableitungen im Affenmodell: Während der Aktivierung des Arbeitsgedächtnisses ist ein Ensemble von Neuronen im präfrontalen Cortex hochgradig aktiv. Der Zusammenhalt dieses Ensembles ist von der Aktivierung von D1-Rezeptoren abhängig. Während der Stimuluspräsentation ist das Neuron moderat aktiv. Verschwindet der Stimulus im Anschluss, ist das Neuron hochgradig aktiv. Das heißt, die aktiven präfrontalen Neurone kodieren wahrscheinlich nicht direkt den gehaltenen Stimulus sondern halten die Aufmerksamkeit auf diejenigen Neuronen im restlichen Gehirn aufrecht, die den relevanten Stimulus verarbeiten. Nachdem die Antwort gegeben wurde, befindet sich das Neuron wieder im niedrig aktiven Ausgangszustand. |
| Kapitel 7: Was ist die Funktion des Hippocampus und welche kognitiven Ausfälle beobachtet man nach Hippocampus-Läsionen? | nicht abgeglichen Aufgabe: - Überführung von Arbeitsgedächtnisinhalten in das Langzeitgedächtnis - Abruf neuerer deklarativer Info - ist wie Adressenverwaltung für cortical abgelegte Erinnerungen - Erinnerungen können aber auch ohne Hippocampus abgerufen werden (Dinge, die häufig erinnert wurden) Hippocampus-Läsionen: - größere Defizite im episodischen als im semantischen Gedächtnis - Nicht deklarative Gedächtnisinhalte sind nicht von Schädigungen des Hippocampus betroffen - beeinträchtigen aber nicht die Intelligenz, die Handlungsfähigkeit und alltägliche kognitive Operationen wie das Sprechen, Addieren oder logische Schlussfolgerungsprozesse. |
| Kapitel 7: Wie funktioniert ein NMDA-Rezeptor? | abgleichen und nochmal lernen NMDA-Rezeptoren brauchen die zeitgleiche Anwesenheit von zwei Ereignissen: die Depolarisation der Zellmembran sowie die Bindung von Glutamat an den NMDA-Rezeptor. Wenn dies passiert (z. B. durch das nahezu zeitgleiche Eintreffen von zwei wichtigen Reizen) wird das Magnesium-Ion, welches den Kanal des NMDA-Rezeptors blockiert, herausgeschleudert und Ca2+-Ionen können in die Zelle einfließen. Sie aktivieren CaM-KII, ein Enzym, welches in dendritischen Spines vorkommt. CaM-KII aktiviert den Einbau von AMPA-Rezeptoren dicht am aktivierten NMDA-Rezeptor. Dadurch wandelt sich die schwache Synapse in eine starke um. In Zukunft können dann auch Einzelreize das Neuron aktivieren, sie viele AMPA-Rezeptoren erregen können. (Beispiel: Nur Hut reicht nicht zur Kanalöffnung, Depolarisation ist zu schwach. Wenn Hut und Kaninchen gesehen werden, löst die zeitliche Summation ein Aktionspotential aus. Depolarisation der NMDA-Membran und Bindung von Glutamat an Rezeptor führt zur Öffnung des Kanals) |
| Kapitel 7: Wie funktioniert ein NMDA-Rezeptor? | Siehe rotes Lernbuch NMDA-Rezeptoren brauchen die zeitgleiche Anwesenheit von zwei Ereignissen: die Depolarisation der Zellmembran sowie die Bindung von Glutamat an den NMDA-Rezeptor. Wenn dies passiert (z. B. durch das nahezu zeitgleiche Eintreffen von zwei wichtigen Reizen) wird das Magnesium-Ion, welches den Kanal des NMDA-Rezeptors blockiert, herausgeschleudert und Ca2+-Ionen können in die Zelle einfließen. Sie aktivieren CaM-KII, ein Enzym, welches in dendritischen Spines vorkommt. CaM-KII aktiviert den Einbau von AMPA-Rezeptoren dicht am aktivierten NMDA-Rezeptor. Dadurch wandelt sich die schwache Synapse in eine starke um. In Zukunft können dann auch Einzelreize das Neuron aktivieren, sie viele AMPA-Rezeptoren erregen können. (Beispiel: Nur Hut reicht nicht zur Kanalöffnung, Depolarisation ist zu schwach. Wenn Hut und Kaninchen gesehen werden, löst die zeitliche Summation ein Aktionspotential aus. Depolarisation der NMDA-Membran und Bindung von Glutamat an Rezeptor führt zur Öffnung des Kanals) |
| Kapitel 7: Wie wird die corticale Konsolidierung während des Schlafs realisiert? Teil 1 | In der zweiten Hälfte mehr REM-Schlaf In der ersten Hälfte: mehr Slow-Wave-Schlaf (langsame EEG-Wellen, ca. 1 Hz) Langsame Wellen entstehen dadurch, dass die Pyramidalneurone des Cortex synchron zwischen einem hochaktiven (up-state) und einem inhibierten (down-state) wechseln Ab und zu kommen aber Ripples vor: hochfrequente hippocampale Oszillationen schnelle neuronale Entladungen, die Ensembles des Cortex und des Hippocampus koppeln Im langsamen Rhythmus des EEG wird mit Hilfe des Hippocampus das neu Gelernte in der synaptischen Struktur des Cortexes stabilisiert Im Maximum einer Slow Wave fangen corticale Neurone an, hochfrequent zu feuern Dadurch werden ihre NMDA-Rezeptoren geöffnet und die beteiligten Synapsen werden verstärkt Gleichzeitig kommt es bei denjenigen Hippocampusneuronen, die im Wachzustand neues gelernt haben, zu Ripples |
| Kapitel 7: Wie wird die corticale Konsolidierung während des Schlafs realisiert? Teil 2 | Das heißt, genau diejenigen Hippocampus-Neurone, die die neue Informationen besitzen, sind nun aktiv, und ihre Aktivität überträgt sich auf die corticalen Zellen, die durch Öffnung ihrer NMDA-Rezeptoren neue synaptische Modifikationen vornehmen und dadurch die hippocampale Information aufnehmen können Der neue Gedächtniseintrag wird so zu einem Teil unseres Gesamtwissens Der Hippocampus lernt schnell, der Cortex langsam In der ersten Hälfte konsolidieren deklarative Gedächtnisinhalte In der zweiten Hälfte konsolidieren prozedurale und emotionale Gedächtnisinhalte |
| Kapitel 7: Was ist der Unterschied zwischen dem Standard-Konsolidierungsmodell und dem multiplen Spurenmodell (Multiple-Trace-Theory)? Oder Wozu brauchen wir den Hippocampus, wenn wir den Cortex haben? | nachholen, steht im Fragenkatalog |
| Kapitel 7: Was ist der Unterschied zwischen Erinnern und Wissen nach dem multiplen Spurenmodell? | In diesem Modell ist „Erinnern“ auf Hippocampus und Cortex angewiesen, während „Wissen“ rein cortical sein kann. Bei einer episodischen Erinnerung kommt es dann zur Aktivierung der gesamten im Hippocampus verknüpften Informationen, sodass auch die zu einer Szene gehörenden Teilkomponenten im Cortex aktiv werden. Bei „Wissen“ wird die semantische Information im Cortex aktiviert, ohne dass die episodischen Verknüpfungen im Hippocampus ebenfalls abgerufen werden. Diese subjektive Unterscheidung zwischen „Wissen“ und „Erinnern“ korreliert in der funktionellen Magnetresonanztomografie mit dem Grad der Aktivierung des Hippocampus. Diese Hippocampusaktivierung ist höher, je subjektiv und objektiv reichhaltiger die episodische Gedächtnisleistung ist ( |
| Kapitel 7: Welche Besonderheiten treten bei frühkindlichen Schädigungen des Hippocampus auf? | Diese Patienten können eine durchaus normale intellektuelle Entwicklung durchmachen und ein durchschnittliches semantisches Gedächtnis entwickeln. Sie haben aber kaum episodische Erinnerungen. Diese Ergebnisse belegen, dass bei sehr früher Schädigung des Hippocampus semantisches Gedächtnis auf corticaler Ebene entstehen kann. Die corticalen Mechanismen können aber bzgl. des episodischen Gedächtnisses nicht den Wegfall des Hippocampus kompensieren. Zudem brauchen diese Patienten mehr Durchgänge mit neuer Information, bis sie diese in ihrem semantischen Gedächtnis gespeichert haben. |
| Kapitel 7: Welche Rolle spielt der präfrontale Cortex beim Abruf von Gedächtnisinformationen? | -PFC sucht Infos, die erinnert werden sollen, aus und koordiniert den Abrufprozess - er kontrolliert beim Abruf zuerst den unbewussten, frühen Kontrollprozess, mit dem wir überprüfen, ob eine Erinnerung überhaupt richtig sein kann - abgerufene Gedächtnisinhalte werden dann vom Frontalcortex editiert, um dann zu entscheiden, ob auf diese Info reagiert werden soll - PFC kann Hippocampus aktiv unterdrücken, um Vergessen zu erzwingen - Eine reduzierte ventrale Aktivierung des PFC führt zu Defiziten im Abruf autobiografischer Information |
| Kapitel 8: Welche nicht deklarativen Gedächtnistypen gibt es und welche Hirnregionen sind für die einzelnen Gedächtnisarten relevant? | - prozedurales Gedächtnis --> Basalganglien - perzeptuelle Bahnung --> sensorische Assoziationsareale - konzeptuelle Bahnung --> PFC und Zugangsstrukturen zum Hippocampus - Klassische Konditionierung --> in Überlappungszonen des CS und des US (bei Lidschlagreaktion ist es z.B. das Kleinhirn) |
| Kapitel 8: Was sind die Hauptunterschiede zwischen prozeduralem und deklarativem Gedächtnis? | Prozedurales Gedächtnis: - schweirig zu beschreiben - kann ohne Beteiligung des Bewusstseins erworben ud ausgeführt werden - braucht Übung - entsteht durch die Interaktion von Cortex und Basalganglien Deklaratives Gedächtnis: - verbal vermittelbar - geht sowohl beim Lernen als auch beim Abrufen mit einer Bewusstseinsbeteiligung einher - entsteht durch die Interaktion von Cortex und Hippocampus |
| Kapitel 8: Welche drei Stufen durchläuft der Konsolidierungsprozess des prozeduralen Gedächtnisses? | 1.) kognitive Stufe: - wir benötigen Instruktion - müssen uns auf Details konzentrieren - Bewusstseinsbeteiligung wird benötigt - Handlung steht unter exekutiver Kontrolle und ist dementsprechen langsam 2.) Assoziative Stufe - Instruktion können wir aus dem deklarativen Gedächtnis abrufen - ansonsten agieren wir wie in der kognitiven Stufe - Sequenz ist gelernt, aber inferenzanfällig - 3.) Autonome Phase - keine Bewusstseinsbeteiligung nötig - automatisierter Ablauf der Handlungen - bewusste Kontrolle kann die Fehlerrate hier sogar erhöhen |
| Kapitel 8: Was sind die neuralen Prozesse, die während des Lernens einer prozeduralen Aufgabe ablaufen? | - Striatumneurone sind bei allen Handlungen involviert - Striatum wird massiv von dopaminergen Fasern innerviert - Freisetzung von Dopamin im Striatum wird reduziert, wenn das Ergebnis der Handlung schlechter war als erwartet - Ist das Ergebnis aber besser als erwartet, erhöht sich die Dopaminfreisetzung im Striatum - Diese sofortige positive oder negative dopaminerge Rückmeldung stärkt die gerade aktiven Synapsen bei Erfolg oder schwächt sie bei Misserfolg -Somit ermöglichen die dopaminergen Rückmeldungen immer bessere Prädiktionen der eigenen Handlungen - Die Zellen des Striatums lernen die erfolgreiche Handlung schneller und fungieren als „Lehrer“ für den Cortex. - Langsam entsteht somit im Kreislauf zwischen Cortex und Basalganglien das Gedächtnis für eine Handlung, die wir von nun an in automatisierter Form abrufen können, während unser Bewusstsein anderen Gedanken nachhängt |
| Kapitel 8: Warum sind Parkinson-Patienten bei prozeduralen Aufgaben gestört? | -Dopaminneurone degenerieren, deren Axone zu den Basalganglien und zum Cortex projizieren - Dadurch kann die Rückmeldung über die Güte der eigenen Handlung an das Striatum nicht mehr erfolgen. Studie von Knowlton et al. (1996) - Wettervorhersage-Aufgabe: --> Symbole auf einer Karte sagen vorher, ob es morgen regnet oder ob die SOnne scheint --> manche Symbole haben eine hohe Vorhersagekraft, manche nicht --> es werden mehrer Karten zeitgleich angeboten --> Karten haben unterschiedliche Vorhersagekraft,Probanden wissen aber nicht, welche wie gut vorhersagen kann --> gesunde Probanden haben Gefühl nur zu raten, aber werden zu ihrem Erstaunen immer besser - Parkinson-Patienten konnten die Aufgaben selbst nach langen Trainingsphasen nicht lernen - Der Ausfall der dopaminergen Rückmeldung an ihre Basalganglein veerhinderte, dass ihre erfolgreiche Striatumneurone das Kommando übernahmen und gleichzeitig die erfolglosen Neurone zum Schweigen verurteilt wurden. Diese Trennung zwischen erfolglosen und erfolgreichen neuronen etablierte sich nicht, deshalb setzte kein lernprozess ein |
| Kapitel 8: Wie funktioniert der Gollin-Figurentest und was passiert in welchen Arealen des Gehirns einer Versuchsperson, die diesen Test durchläuft? | Beim Gollin-Figurentest wird ein immer weniger fragmentierter Stimulus mehrfach angeboten bis sich eine Gedächtnisspur für diesen Reiz etabliert hat. Nach einer längeren Pause wiederholt man den Versuch. Diesmal wird typischerweise beobachtet, dass die Versuchspersonen die gesuchten Gegenstände in einer wesentlich früheren und somit unvollständigeren Version erkennen können. Zu Beginn sind sehr viele Zellen für dieses Objekt aktiv. Diese stärken sich mit der Zeit, sodass am Ende eine “Expertengruppe” übrig bleibt (sparsames Kodieren). Das heißt, es hat sich ein perzeptuelles Gedächtnis für diese Objekte gebildet. Dieses Gedächtnis bildet sich hauptsächlich in den assoziativen sensorischen corticalen Arealen und ist hippocampus-unabhängig. |
| Kapitel 8: Wie funktioniert konzeptuelle Bahnung und welche Hirnregionen sind relevant? | Konzeptuelle Bahnung = Die größere Wahrscheinlichkeit, ein Objekt einer bestimmten Kategorie zu erkennen oder zu erinnern, wenn vorher diese Kategorie präsentiert wurde. Es wird eine Begriffskategorie aktiviert. Nach einer Weile erkennen Versuchspersonen eher Objekte, die zu dieser Kategorie gehören. Die kozeptuelle Bahnung ist wesentlich modalitätsunabhängiger als die perzeptuelle Bahnung. Kritische Hirnregionen sind der (linke) Präfrontalcortex und die Zugangsregionen zum Hippocampus (medialer Temporallappen). Alzheimer-Patienten zeigen daher zwar keine Defizite in der perzeptuellen bahnung, aber dafür in der konzeptuellen Bahnung. |
| Kapitel 8: Erläutern Sie die Hirnmechanismen der klassischen Konditionierung am Beispiel der Lidschlagkonditionierung. | nochmal intensiv bearbeiten |
| Kapitel 9: Definieren sie die Eigenschaften und Funktionen von Emotionen aus biopsychologischer Sicht. | - Emotionen sind Handlungstendenzen - gehen mit einem bestimmten Gefühl sowie der gebündelten Aktivierung eines Pakets von Hirnstrukturen und Hormonsystemen einher - Diese Prozesse bereiten den Organismus auf eine schnelle und situationsadäquate Reaktion vor - Gefühle sind die subjektive Innenansicht einer Emotion, die uns auf eine Handlung vorbereitet - Emotionen entstanden evolutionär aus der Notwendigkeit, möglichst schnell ein koordiniertes Bündel motorischer Reaktionen zur Hin- oder Abwendung von einem Stimulus oder einer Situation zu aktivieren. |
| Kapitel 9: Was ist das limbische System und welche Anteile des Gehirns gehören dazu? | - organisiert emotional moduliertes Verhalten und wurde als eine Gruppe von Vorderhirnregionen definiert, die alle mit dem Hippocampus verbunden sind - es werden je nach Autor nicht immer die gleichen Hirnregionen aufgezählt - diese Regionen sind aber eig immer involviert: Amygdala, Septum, Hippocampus, anteriore Kerne des Thalamus, Hypothalamus, Mammilarkörper, cingulärer Cortex, OFC |
| Kapitel 9: Wie ist die Amygdala aufgebaut und was sind ihre relevanten Konnektivitäten? | - besteht bei Primaten aus 13 Unterkernen, die in 3 große Gruppen aufgeteilt werden: 1.) basolateral 2.) centromedial 3.) corticomedial - lateraler Amygdalakern (LA, basolateral) ist das sensorische Einfallstor der Amygdala --> erhält Eingänge aus Cortex, Thalamus und innerhalb der basolateralen Kerngruppe zum basalen Kern der Amygdala (BA) - BA-Neurone haben reziproke Verbindungen zu vielen cortikalen Arealen sowie indirekt zum Hippocampus - diese Verbindungen beeinflussen die emotionale Modulation der Gedächtnisbildung - BA projiziert zum zentralen Kern der Amygdala (Ce) - Ce projiziert in den Hypothalamus und weiten Teilen des Hirnstamms und aktivert angeborene Furchreaktionsmechanismne - Medialer kern (Me) gehört zur corticomeialen Gruppe der Amygdalakerne, erhält versch. sens. Eingänge, wobei der Input aus dem olfaktorischen System dominert - Me hat reziproke Verbindungen zum medialen Hypothalamus und spielt eine wichtige Rolle bei der Kontrolle des Sexual- und Aggressionsverhaltens |
| Kapitel 9: Erläutern Sie anatomisch, warum Katzen nicht gleichzeitig auf Mäusefang gehen und mit einem Artgenossen kämpfen können. | - ventrales Höhlengrau kann motorische Prozesse des Beutefangs aktivieren - Teile des lateralen Hypothalamis (hier werden Hungersignale verarbeitet) kontrollieren das ventrale Höhlengrau und somit das instrumentelle Beuteverhalten - Aggression stammt aus dorsalem Höhlengrau/ medialer Hypothalamus - medialer und lateraler Hypothalamus hemmen sich gegenseitig --> aggressive Handlungen und Nahrungsaufnahme sind somit unvereinbar - zentraler Kern der Amyggdala (CA) hemmt das dorsale Höhlengrau --> Angst und Fluchtverhalten (Ce) sind nicht mit Aggressionsverhalten kompatibel. --> kann diese beiden Dinge nie gleichzeitig tun |
| Kapitel 9: Beschreiben Sie die high road und die low road der Erkennung emotionaler Reize. | High road: Retina --> Thalamus --> Sehrinde --> Amygdala Low road: Retina --> Thalamus --> Amygdala Die Amygdala bekommt sensorische Eingänge aus thalamischen Strukturen, die mehrere Reizmodalitäten integrieren. Diese Kerne vermitteln nur eine sehr grobkörnige Repräsentation der Außenwelt, aber dafür erfolgt ihre Reizverarbeitung sehr schnell. Das bedeutet, dass die Amygdala sehr früh einen groben Informationsfluss von der sensorischen Umwelt erhält („low road“). Wenn sich in unserer Umwelt etwas potenziell Gefährliches verbirgt, kann die Angstmaschinerie der Amygdala sehr schnell angeworfen werden und eine Reaktion erfolgt in kürzester Zeit. Dagegen braucht die Verarbeitung derselben Reize durch die corticalen Mechanismen länger, aber sie erzeugt dafür ein feinkörniges und uns bewusst zugängliches Abbild der Welt („high road“). |
| Kapitel 9: Führen Läsionen der Amygdala zur Unfähigkeit emotionale Reize zu erkennen? | Nein. Amygdala-Läsionen führen dazu, dass die emotional relevanten Regionen eines Gesichtes nicht beachtet werden. Wenn man die Patienten auffordert, diese Regionen zu beachten, sind sie durchaus in der Lage, Auskunft über die richtige Emotion zu geben. Das heißt, man ist ohne Amygdala nicht “angstblind”, sondern man weiß nur nicht, wo in der Umwelt die diagnostischen Hinweisreize zu suchen sind. |
| Kapitel 9: Was sind die Mechanismen, mit denen wir bei emotionalen Abläufen die Hauptdetails des Geschehens besonders gut erinnern? | Der basale Kern der Amygdala (BA) hat reziproke Verbindungen zum Hippocampus und kann somit semantische und episodische Gedächtnisinhalte modifizieren. Zudem gibt es eine Projektion des zentralen Kerns der Amygdala (Ce) auf den Nucleus basalis von Meynert (im Frontalcortex), der für die Acetylcholin-Ausschüttung im Cortex relevant ist. Diese Acetylcholimpulse treten immer dann auf, wenn ein emotionaler Reiz auftritt. Dadurch werden Umgebungsreize, die im Moment dieses Reizes vorlagen und somit gerade im Cortex repräsentiert wurden mit der Emotion verbunden. Tatsächlich zeigen Patienten mit Amygdala-Läsionen keine Fähigkeit mehr, die emotional getönten Aspekte eines Ereignisses ganz besonders gut zu erinnern. |
| Kapitel 9: Wie funktioniert Furchtkonditionierung auf zellulärem Niveau in der Amygdala? | nochmal im Echo anhören und mit Buch und Skript abgleichen --> Buch Seite 289 Der eigentliche Lernprozess der Furchtkonditionierung beginnt im lateralen Amygdalakern. Da hier durch die Multimodalität des thalamischen Inputs alle sensorischen Aspekte eines emotionalen Reizes auf einzelnen Neuronen konvergieren, kommt es zu einer Veränderung der Antworteigenschaften der Neurone entsprechend der Hebb'schen Regel. Diese Regel besagt, dass schwache Reize (konditionierter Stimulus; CS) eine zunehmend stärkere synaptische Wirkung entwickeln, wenn sie immer wieder mit einem starken Reiz (unkonditionierter Stimulus; UCS) gepaart werden. Die Zellen des LA reagieren somit nach mehreren CS-UCS-Paarungen auf den CS alleine, auch wenn er nicht mehr vom UCS gefolgt wird. Über die Projektion des LA zum basalen Kern der Amygdala (BA) und von dort zum zentralen Kern der Amygdala (Ce) kommt es nun nach CS-Vergabe zu den mit Furcht assoziierten Verhaltensreaktionen. |
| Kapitel 10: Wie kann es sein, dass, obwohl die Wirkung aller Drogen auf der Aktivierung der mesolimbischen Bahn beruht, sich dennoch die Details der Drogenmechanismen unterscheiden? | nicht abgeglichen Die Einnahme von Drogen führt zu Erhöhung des Dopaminspiegels im Nucleus accumbens und erzeugt einen Lustgewinn. Dies liegt daran, dass viele Drogen die Dopaminneurone des ventralen tegmentalen Areals (VTA) aktivieren können, somit eine Dopaminfreisetzung im Nucleus accumbens bewirken und dadurch dieses Areal aktivieren. Aber die Neuronen des Nucleus accumbens haben auch Rezeptoren für Opioide, Alkohol, Nikotin und Cannabis. Das heißt, dass die Zellen des Nucleus accumbens von diesen Substanzen auch direkt aktiviert werden können. Dadurch sind diese Substanzen von Läsionen der mesolimbischen Dopaminbahn unabhängiger. Durch diese Mischung aus Rezeptoren in der VTA bzw. im Nucleus accumbens gibt es für verschiedene Stoffe unterschiedliche Wege, den Nucleus accumbens zu aktivieren. |
| Kapitel 10: Beschreiben Sie die Opponent-Prozess-Theorie von Solomon und Corbitt und führen Sie aus, wie sich das Ergebnis von a- und b-Prozessen durch den Grad der Gewöhnung verändern. | Jede Emotion , aber auch die Einnahme einer Droge, erzeugt zwei gegensätzliche Effekte, den a-Prozess und den b-Prozess. Der a-Prozess ist hedonische Resultat der Droge und bildet Intensität und Wirkdauer ab. b-prozess ist zeitverzögert und weniger intensiv. (die anfängliche Freude zieht ein bisschen Trauer nach sich). Der b-Prozess ist schwächer als der a-Prozess, aber seine Stärke korreliert mit dem a-Prozess. Was wir subjektiv erleben, ist die Summation des a- und des b-Prozesses. Wenn wir uns aber an Droge gewöhnen, wird der b-Prozess stärker, beginnt schneller, bleibt aber trotzdem immer ein bisschen kleiner als der a-Prozess. Die Summation der beiden Prozesse sieht nun so aus, dass der anfängliche Euphoriegipfel kleiner ist, das Plateau nahe der Nulllinie verläuft und das depressive Ende stärker ist als der Euphoriegipfel. Um die Euphorie wieder herzustellen, muss die Drogenmenge erhöht werden (schleichende Erhöhung der konsumierten Drogenmenge). Zusätzlich werden die Drogen häufiger eingenommen, um der negativen Nachschwankung des b-Prozesses entgegenzuwirken. |
| Kapitel 10: Wie und wo entstehen die Netzwerke der Drogenkonsum-assoziierten Hinweisreize und Handlungen im Gehirn? | Dafür sind 2 Aspekt wichtig: 1.) Bei wiederholtem Konsum einer Droge in best. Situationen läuft dopaminerge Freisetzung zeitlich „rückwärts“. D.h. Dopamin wird immer zum Zeitpunkt des nächstfrüheren Prädiktors des Drogenkonsums freigesetzt. Am Ende wird Dopamin freigesetzt, wenn Handlungskette angestoßen wird, an deren Ende die Droge steht. --> z.B. in die Tasche greifen 2.) Diese Handlungsketten haben verschiedene Ebene der Kontrolle durch das Gehirn. --> Niedrigste Ebene: motorisch (motorischer Cortex, dorsales Striatum) --> es entstehen Bewegungsmuster, die wir ohne Nachdenken ausführen (z. B. beiläufig in die Tasche greifen) --> mittlere Ebene: hippocampale Verarbeitung und Erkennung eines Kontextes, welches mit Zigarettenkonsum assoziiert ist (z. B. Geselligkeit). --> höchste Stufe: präfrontale Handlungsplanung, bei der wir Situation aufsuchen, in der Rauchen wahrscheinlich ist (z. B. sich mit Freunden zu treffen) Solche situativen Merkmale sind am Ende so etabliert, dass sie Leerlauffunktionen übernehmen. Wenn uns lw ist, suchen wir Zigaretten-Situationen auf |
| Kapitel 10: Beschreiben Sie die synaptischen Zusammenhänge im VTA (ventralen tegmentalen Areal) am Beispiel der Nikotinsucht. | nochmal in ruhe anschauen und ins lernbuch packen |
| Kapitel 10: Wieso feuert ein dopaminerges Neuron schon beim Brötchenschmieren, obwohl noch gar keine Zigarette in Sicht ist? | Dies passiert nur, wenn das Schmieren des Brötchens das erste Glied einer Kette ist, an deren Ende das Rauchen einer Zigarette steht. In einem solchen Fall feuerte das dopaminerge Neuron ganz am Anfang der Etablierung einer Verhaltenskette beim Rauchen der Zigarette. Je häufiger eine Prozedur wiederholt wird, desto mehr wird das Dopamin zu früheren Zeitpunkten ausgeschüttet, die den jeweils nächsten konditionierten Stimulus vorhersagen können (Greifen nach der Packung, Trinken des letzten Schluck Kaffee, Abbeißen vom Brötchen etc.) bis es letztendlich beim allerersten konditionierten Reiz angekommen war, nämlich dem Schmieren des Brötchens. |
| Kapitel 10: Welche Prozesse folgen der Abstinenz? | nicht abgeglichen Bei Abstinenz sinkt der Dopaminspiegel im Nucleus accumbens und es entsteht Reizbarkeit, Schreckhaftigkeit und sogar leichte kognitive Defizite. Es entsteht auch eine gedrückte Stimmung. Dazu kommt das Freisetzen von Stresshormonen und somit eine Erhöhung des Angstniveaus selbst bei harmlosen Situationen. Solche Situationen werden häufig dann auch als eher unangenehm erlebt. |
| Kapitel 11: Wie speichern wir die Energie aus der Nahrung? | - Im Darm wird Nahrung in Kohlenhydrate, Glucose, Fette und Aminosäuren zerlegt - Anstieg von Glucose im Blut (= Blutzucker) ist das Signal für die Erzeugung von INsulin durch die Pankreas. Dadurch können Körperzellen Glucose aufnehmen -Glucose = Energielieferant für Zellen - Gehirn verbraucht 50 Prozent der Glucose, da Natrium-Kalium-Pumpe hohen Energieverbrauch hat und kann auch ohne Insulin Glucose aufnehmen --> Gehirn = "egoistisches Organ" Für kurzfristige Speicherung wird Glucose mithilfe von Insulin in Glykogen verwandelt. Glykogen kann in der Leber und in den Muskeln gespeichert werden. Weitere Nährstoffe werden in Triglyceride umgewandelt und in den Fettzellen des Fettgewebes gespeichert., sie sich enorm vergößern können. Dies ist unser langfristiger Speicher. |
| Kapitel 11: Welche Rolle spielen die Hormone Leptin und Ghrelin bei der Regulation des Essverhaltens? | siehe Word |
| Kapitel 11: Wie wirkt Vasopressin und welche Prozesse führen zu seiner Freisetzung? | nicht abgelgichen - Vasopressin verändert die Filtereigenschaften der Niere, sodass vermehrt Wasser aus dem Harn zurückgewonnen wird. - Dadurch verdickt sich der Urin und weniger Wasser geht verloren. - Vasopressin wird nach einem Absinken des osmometrischen Wasserdrucks freigesetzt und zwar durch die Aktivierung des Nucleus paraventricularis und des Nucleus supraopticus. - Beide Strukturen produzieren Vasopressin und transportieren es in den hinteren Teil der Hypophyse. |
| Kapitel 11: Was unterscheidet osmometrischen von volumetrischem Durst? | nicht abgeglichen Osmometrischer Durst entsteht dadurch, dass durch Wasserverlust der relative Anteil der Ionen in der interstiziellen Flüssigkeit (Flüssigkeit im Interzellularraum) ansteigt. --> Da diese unsere Körperzellen umfließt, entsteht ein osmotischer Druck, der den Körperzellen Wasser entzieht und diese schrumpfen. Volumetrischer Durst entsteht durch Blutverlust. Dadurch sinkt der Blutdruck und dies wird von Dehnungssensoren (Barorezeptoren) in den Vorhöfen unseres Herzens registriert. Bei einem Blutverlust fließt auch weniger Blut durch die Nieren. Dies wird durch Sensoren registriert, die Blutflussmenge messen. |
| Kapitel 11: Wie erzeugt osmometrischer Durst Trinkverhalten? | nicht abgeglichen Das Schrumpfen der Zellmembran wird vom Organum vasculosum der Lamina terminalis (OVLT) und dem Subfornikalorgan (SFO) detektiert. Beide projizieren zum medialen präoptischen Nukleus (MPO), der wiederum im lateralen Hypothalamus die Orexin-Neurone aktiviert. Diese Zellen lösen im Cortex kognitive Prozesse des Trinkens an. OVLT & SFO MPO Orexin-Neurone im lateralen Hypothalamus Cortex Trinken |
| Kapitel 11: Wie erzeugt volumetrischer Durst Trinkverhalten? | nicht abgeglichen Die Barorezeptoren schlagen bei Blutdruckabfall Alarm und senden ihre Signale durch den Nervus vagus zum Nucleus solitarius der Medulla oblongata. Der Nucleus solitarius projiziert über die präoptische Region zu den Orexin-Neuronen im lateralen Hypothalamus, die das Trinken kontrollieren. Bei einem Blutverlust fließt auch weniger Blut durch die Nieren. Daraufhin setzt die Niere das Enzym Renin frei. Dieses gelangt ins Blut und katalysiert dort über Angiotensinogen Angiotensin und schließlich Angiotensin II. Die Zellen des Subfornikalorgans besitzen Rezeptoren für Angiotensin II und können somit über die für das volumetrische System beschriebenen Mechanismen Trinkverhalten auslösen. Barorezeptoren --> Nervus vagus --> Nucleus solitarius der Medulla Oblongata --> über präoptische Region zu Orexin-Neuronen im lateralen Hypothalamus --> Niere setzt Renin frei --> Renin ins Blut und katalysiert dort über Angiotensinogen Angiotensin und Angiotensin II --> Zellen des Subfornikalorgans besitzen Rezeptoren für Angiotensin II und können so Trinkverhalten auslösen |
| Kapitel 11: 6a) Welche besonderen Eigenschaften weisen circumventriculäre Organe auf? | nicht abgelgichen - unpaarige Nuklei, die entlang der Medianebene des Gehirns direkt am Ventrikel liegen. - In ihnen ist die Blut-Hirn-Schranke aufgehoben. Das heißt, alle im Blut gelösten Stoffe können die Neurone der circumventrikulären Organe direkt beeinflussen. - Stattdessen besitzen diese Nuklei eine Ventrikel-Hirn-Schranke, sodass die in der Ventrikelflüssigkeit gelösten Hormone und Transmitter nicht direkt die Zellen der circumventrikulären Organe beeinflussen können. |
| Kapitel 11: 6b) Wieso ist diese Eigenschaft für die Wirkung von Angiotensin II bedeutsam? | nicht abgeglichen Die Zellen des Subfornikalorgans besitzen Rezeptoren für Angiotensin II und können dieses Hormon direkt aus dem Blut aufnehmen. |
| Kapitel 11: Wie aktiviert die Reduktion unserer Energiereserven nahrungsbezogenes Verhalten? I | - Bei Reduktion unserer Energiereserven kommt es zum Absinken des Blutzuckerspiegels - In der Medulla oblongata sitzen Rezeptoren für Glucose und kontrollieren die Bewegungsabläufe der Nahrungsaufnahme - In diesem Bereich enden auch Fasern des Vagusnerven, der das Gehirn über den Abfall des Fettspiegels in der Leber und über Aktivierung von Nährstoffrezeptoren im Magen und im Zwölffingerdarm informiert. Dadurch aktiviert Medulla oblongata einen Teil des nahrungsbezogenen Verhaltens. |
| Kapitel 11: Wie aktiviert die Reduktion unserer Energiereserven nahrungsbezogenes Verhalten? II | - Wenn Fettzellen mit genügend Triglyceriden gefüllt sind, produzieren sie Leptin - Das Sinken des Leptinspiegels signalisiert den Zellen des Nucleus arcuatus im Hypothalamus, dass neue Nahrung gebraucht wird - Dadurch wird in diesem Hirnkern ein Neuronentyp aktiviert, der NPY und AGRP als Neurotransmitter verwendet - Diese Zellen erkennen noch ein weiteres Hormon, Ghrelin, das vom leeren Magen freigesetzt wird. - Die Axone der NPY- und AGRP-Neurone terminieren in zwei benachbarten Arealen, dem PVN und dem lateralen Hypothalamus - Im PVN hemmen die Fasern aus dem Nucleus arcuatus die Freisetzung von ACTH und TSH - ACTH und TSH steigern den Stoffwechsel & erhöhen die Körpertemperatur - Hemmung dieser Hormone lässt den Körper somit auf Sparflamme laufen und minimiert unnötigen Energieverbrauch - Die Projektion der NPY- und AGRP-Zellen auf lateralen Hypothalamus hat dagegen einen ganz anderen Effekt: Dort werden Neurone aktiviert, die MCH und Orexin freisetzen. - Die MCH- und Orexin-Zellen haben Axone, die in den Cortex reichen und dort Nahrungssuche initiieren. |
| Kapitel 11: Wie aktiviert die Reduktion unserer Energiereserven nahrungsbezogenes Verhalten? III | - Ein anderer Teil projiziert in die AVT und aktiviert die Dopaminfreisetzung, wodurch die Erwartung von Belohnung durch die Nahrungsaufnahme entsteht. - Wieder andere MCH- und Orexin-Axone führen zu einer allgemeinen Aktivierung des Gehirns. Diese erhöhte Wachheit unterstützt die Nahrungssuche. |
{kind=link}
¿Quieres crear tus propias Fichas gratiscon GoConqr? Más información.