18781108
Description
Flashcards by Diana Rivera, updated more than 1 year ago
|
|
Created by Diana Rivera
over 5 years ago
|
|
| Question | Answer |
| Las funciones reproductoras masculinas pueden dividirse en tres apartados principales: | 1) la espermatogenia, que significa la formación de los espermatozoides; 2) la realización del acto sexual masculino, y 3) la regulación de las funciones reproductoras del varón por diversas hormonas. Asociados a estas funciones reproductoras están los efectos de las hormonas sexuales masculinas en los órganos sexuales accesorios, el metabolismo celular, el crecimiento y otras funciones del organismo. |
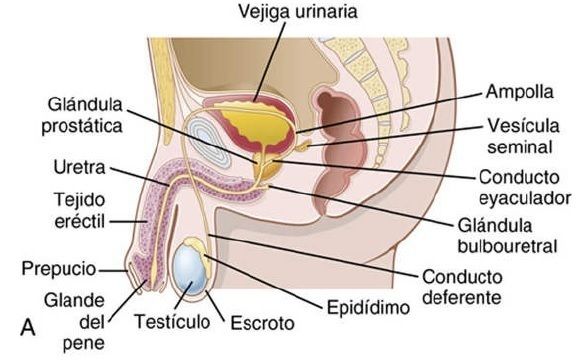
| *Anatomía fisiológica de los órganos sexuales masculinos* La figura 81-1A muestra las distintas partes del aparato reproductor masculino |
Image:
Az (binary/octet-stream)
|
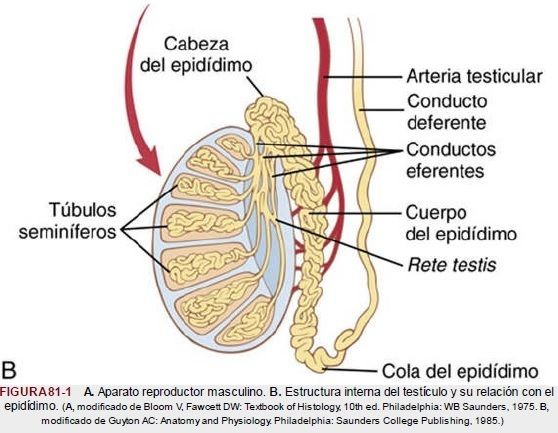
| La estructura del testículo y del epidídimo |
Image:
Am (binary/octet-stream)
|
| El testículo | está compuesto por hasta 900 túbulos seminíferos espirales, cada uno de más de 0,5 m de longitud, en los que se forman los espermatozoides. Estos se vacían después al epidídimo, que es otro tubo espiral de unos 6 m de longitud. El epidídimo se abre al conducto deferente, que se ensancha para formar la ampolla del conducto deferente inmediatamente antes de su desembocadura en el cuerpo de la glándula prostática. |
| Dos vesículas seminales, localizadas una a cada lado de la próstata, desembocan en el extremo prostático de la ampolla y el contenido de esta y de las vesículas seminales pasa al conducto eyaculador, que atraviesa el cuerpo de la glándula prostática para finalizar en la uretra interna. | Los conductos prostáticos van desde la próstata al conducto eyaculador y desde él a la uretra prostática. Por último, la uretra es el eslabón final de la comunicación del testículo con el exterior. La uretra está provista de moco procedente de numerosas glándulas uretrales diminutas localizadas en toda su longitud y, todavía en mayor cantidad, de las glándulas bulbouretrales (glándulas de Cowper) bilaterales situadas cerca del origen de la uretra. |
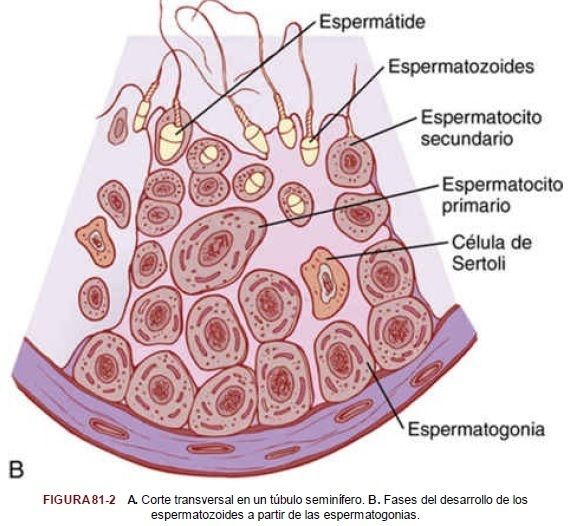
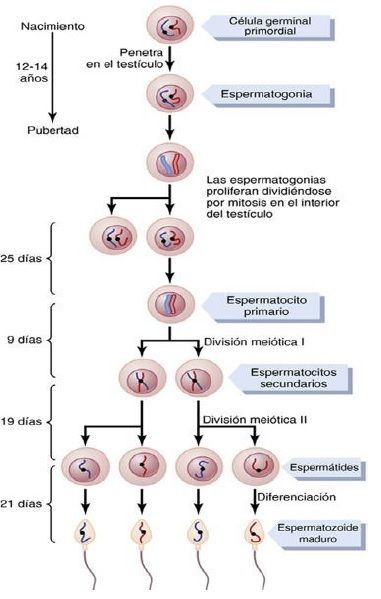
| *Espermatogenia* Durante la formación del embrión, las células germinales primordiales migran hacia los testículos y se convierten en células germinales inmaduras llamadas espermatogonias, que ocupan las dos o tres capas más internas de los túbulos seminíferos. FIGURA 81-2 A. Corte transversal en un túbulo seminífero. |
Image:
An (binary/octet-stream)
|
| Como aparece en la figura 81-2B, a partir de la pubertad las espermatogonias comienzan a dividirse por mitosis y continúan proliferando y diferenciándose a los estadios definitivos de desarrollo para formar espermatozoides. |
Image:
Ad (binary/octet-stream)
|
| *Pasos de la espermatogenia* La espermatogenia tiene lugar en todos los túbulos seminíferos durante la vida sexual activa, como consecuencia de la estimulación por las hormonas gonadótropas de la adenohipófisis, comenzando por término medio a los 13 años y continuando durante el resto de la vida, aunque disminuye notablemente en la vejez. | En esta primera fase, las espermatogonias migran hacia la luz central del túbulo seminífero entre las células de Sertoli. Las células de Sertoli son muy grandes, con cubiertas de citoplasma redundantes que rodean a las espermatogonias en desarrollo hasta la luz central del túbulo. |
| *Meiosis* Las espermatogonias que atraviesan la barrera y penetran en la capa de células de Sertoli se modifican progresivamente y aumentan de tamaño para formar espermatocitos primarios grandes (fig. 81-3). Cada espermatocito primario se divide para formar dos espermatocitos secundarios. Al cabo de unos pocos días, estos espermatocitos se dividen a su vez para formar espermátides, que tras varias modificaciones acaban convirtiéndose en espermatozoides (esperma). |
Image:
Af (binary/octet-stream)
|
| Durante la etapa de modificación desde la fase de espermatocito a la de espermátide, los 46 cromosomas (23 pares de cromosomas) del espermatocito se reparten, de manera que: | 23 cromosomas van a una espermátide y los otros 23, a la otra. Esto también hace que se dividan los genes cromosómicos, de manera que solo una mitad del material genético de un posible feto procede del padre y la otra mitad procede del ovocito de la madre. |
| Todo el período de espermatogenia, desde la espermatogonia hasta el espermatozoide, tiene una duración aproximada de | 74 días. |
| *Cromosomas sexuales* En cada espermatogonia, uno de los 23 pares de cromosomas transporta la información genética que determina el sexo del descendiente. Este par está compuesto por un cromosoma X, denominado cromosoma femenino, y un cromosoma Y, el cromosoma masculino. | Durante la división meiótica, el cromosoma masculino Y se dirige a una espermátide, que se convierte en un espermatozoide masculino, y el cromosoma femenino X va a otra espermátide, que se convierte en un espermatozoide femenino. El sexo de la descendencia dependerá de cuál de estos dos tipos de espermatozoides fecunde al óvulo. |
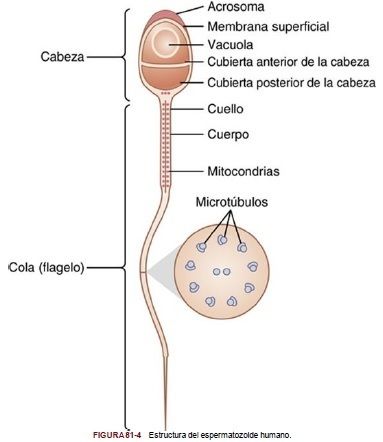
| *Formación del espermatozoide* Cuando las espermátides se forman por primera vez, tienen todavía las características habituales de las células epitelioides, pero pronto cada espermátide comienza a alargarse para constituir los |
espermatozoides, cada uno compuesto por cabeza y cola.
Image:
Es (binary/octet-stream)
|
| La cabeza está formada por el núcleo celular condensado revestido tan solo de una fina capa de citoplasma y de membrana celular en torno a su superficie. En la parte externa de los dos tercios anteriores de la cabeza existe una capa gruesa denominada | acrosoma, consistente sobre todo en el aparato de Golgi. El acrosoma contiene varias enzimas similares a las que se encuentran en los lisosomas de las células típicas, incluida la hialuronidasa (que puede digerir los filamentos de proteoglucanos de los tejidos) y poderosas enzimas proteolíticas (que pueden digerir proteínas). Estas enzimas desempeñan funciones importantes, pues permiten al espermatozoide entrar en el óvulo y fecundarlo. |
| La cola del espermatozoide, denominada flagelo, tiene tres componentes principales: | 1) un esqueleto central constituido por 11 microtúbulos, denominados en conjunto axonema, cuya estructura es similar a la de los cilios de las superficies de otros tipos de células, 2) una fina membrana celular que reviste el axonema, y 3) una serie de mitocondrias que rodean el axonema de la porción proximal de la cola (denominada cuerpo de la cola). |
| El movimiento de vaivén de la cola (movimiento flagelar) determina la motilidad del espermatozoide. Este movimiento es el resultado de un movimiento rítmico de deslizamiento longitudinal entre los túbulos anteriores y posteriores que constituyen el axonema. La energía necesaria para este proceso procede del | trifosfato de adenosina sintetizado por las mitocondrias del cuerpo de la cola. |
| Los espermatozoides normales se mueven en medio líquido a una velocidad de | 1 a 4 mm/min, lo que les permite desplazarse a través del aparato genital femenino en busca del óvulo. |
| Factores hormonales que estimulan la espermatogenia. He aquí algunas de ellas: | 1. La testosterona, 2. La hormona luteinizante, 3. La hormona foliculoestimulante, 4. Los estrógenos, 5. La hormona del crecimiento |
| 1. La testosterona, | secretada por las células de Leydig localizadas en el intersticio testicular, es esencial para el crecimiento y la división de las células germinales testiculares, que es el primer paso en la formación de los espermatozoides. |
| 2. La hormona luteinizante, | secretada por la adenohipófisis, estimula la secreción de testosterona por las células de Leydig. |
| 3. La hormona foliculoestimulante, | también secretada por la adenohipófisis, estimula a las células de Sertoli; sin esta estimulación no se produciría la conversión de espermátides en espermatozoides (el proceso de la espermatogenia). |
| 4. Los estrógenos, | formados a partir de la testosterona por las células de Sertoli cuando son estimuladas por la hormona foliculoestimulante, también son, probablemente, esenciales para la espermatogenia. |
| 5. La hormona del crecimiento (al igual que la mayoría de las restantes hormonas) es necesaria para | controlar las funciones metabólicas básicas de los testículos. En concreto, la hormona del crecimiento promueve la división temprana de las propias espermatogonias; en su ausencia, como ocurre en el enanismo hipofisario, la espermatogenia es muy deficiente o nula, lo que se traduce en esterilidad. |
| *Maduración del espermatozoide en el epidídimo* Tras su formación en los túbulos seminíferos, los espermatozoides tardan varios días en recorrer el epidídimo, un tubo de | 6 m de largo. |
| Los espermatozoides extraídos de los túbulos seminíferos y de las primeras porciones del epidídimo son inmóviles e incapaces de fecundar un óvulo. Sin embargo, tras haber permanecido en el epidídimo entre 18 y 24 h, desarrollan la capacidad de | motilidad, aunque diversas proteínas inhibidoras del líquido del epidídimo impiden el movimiento real hasta después de la eyaculación. |
| *Almacenamiento de los espermatozoides en los testículos* Los dos testículos del ser humano adulto forman unos | 120 millones de espermatozoides diarios. |
| La mayoría de los espermatozoides se conservan en el conducto deferente, aunque en una pequeña cantidad se almacenan en el epidídimo. Pueden permanecer almacenados, manteniendo su fertilidad, durante al menos 1 mes. | En este tiempo se mantienen en un estado de profunda inhibición provocado por múltiples sustancias inhibidoras de las secreciones de los conductos. Por el contrario, con una actividad sexual y eyaculaciones excesivas, el almacenamiento a veces no dura más de unos pocos días a lo sumo. |
| Tras la eyaculación, los espermatozoides se vuelven móviles y capaces de fecundar al óvulo, un proceso denominado | maduración. Las células de Sertoli y el epitelio del epidídimo secretan un líquido nutritivo especial que es eyaculado junto con los espermatozoides. Este líquido contiene hormonas (testosterona y estrógenos), enzimas y nutrientes especiales, imprescindibles para la maduración de los espermatozoides. |
| *Fisiología del espermatozoide maduro* Los espermatozoides normales, móviles y fértiles, son capaces de movimientos flagelares a través de un medio líquido a una velocidad de | 1 a 4 mm/min. La actividad de los espermatozoides es mucho más fácil en el medio neutro y algo alcalino del semen eyaculado, pero se deprime mucho en los medios ligeramente ácidos. Los medios muy ácidos provocan la muerte rápida de los espermatozoides. |
| La actividad de los espermatozoides aumenta notablemente a medida que se eleva la temperatura, pero también lo hace su metabolismo, lo que acorta de manera considerable su supervivencia. Aunque los espermatozoides pueden sobrevivir muchas semanas en los conductos genitales de los testículos, su supervivencia en el aparato genital femenino es de solo | 1 o 2 días. |
| *Función de las vesículas seminales* Cada vesícula seminal es un túbulo tortuoso, lobulado, revestido por un epitelio secretor que genera un material mucoide rico en | fructosa, ácido cítrico y otras sustancias nutritivas, así como grandes cantidades de prostaglandinas y fibrinógeno. |
| Durante el proceso de emisión y eyaculación, cada vesícula seminal vacía su contenido al | conducto eyaculador poco tiempo después de que el conducto deferente libere los espermatozoides. Esta contribución aumenta mucho el volumen de semen eyaculado y la fructosa y otras sustancias del líquido seminal tienen un considerable valor nutritivo para los espermatozoides eyaculados, hasta que uno de ellos fecunda el óvulo. |
| Se cree que las prostaglandinas ayudan de dos maneras a la fecundación: | 1) reaccionando con el moco cervical femenino, para hacerlo más receptivo al movimiento de los espermatozoides, y 2) posiblemente, desencadenando contracciones peristálticas invertidas del útero y de las trompas de Falopio para desplazar a los espermatozoides hacia los ovarios (unos pocos espermatozoides alcanzan el extremo superior de las trompas de Falopio en 5 min). |
| *Función de la próstata* | La próstata secreta un líquido poco denso, lechoso, que contiene iones citrato, calcio y fosfato, una enzima de coagulación y una profibrinolisina. Durante la emisión, la cápsula de la próstata se contrae en paralelo con las contracciones del conducto deferente, de forma que el líquido poco denso y lechoso de la próstata contribuye aún más al volumen de semen. |
| El carácter ligeramente alcalino de este líquido podría ser bastante importante para el éxito de la fecundación del óvulo, pues el líquido del conducto deferente es relativamente ácido por la presencia del ácido cítrico y de los productos finales del metabolismo de los espermatozoides y, en consecuencia, ayuda a inhibir la fertilidad de los espermatozoides. | Además, las secreciones vaginales de la mujer son ácidas (con un pH de 3,5 a 4). Los espermatozoides no alcanzan una motilidad óptima hasta que el pH del líquido que los baña se eleva de 6 a 6,5. En consecuencia, es probable que el líquido prostático, algo alcalino, ayude a neutralizar la acidez de estos otros líquidos tras la eyaculación y facilite la movilidad y fertilidad de los espermatozoides. |
| *Semen* El semen, eyaculado durante el acto sexual masculino, se compone del | líquido y los espermatozoides del conducto deferente (aprox. el 10% del total), el líquido de las vesículas seminales (aprox. el 60%), el líquido de la glándula prostática (aprox. el 30%) y pequeñas cantidades procedentes de las glándulas mucosas, sobre todo de las glándulas bulbouretrales.Por tanto, el grueso del volumen del semen es líquido de las vesículas seminales, que es el último en ser eyaculado y sirve para lavar los espermatozoides del conducto eyaculador y la uretra. |
| El pH medio del semen mezclado es de alrededor de | 7,5, pues el líquido prostático alcalino neutraliza la ligera acidez de las otras porciones del semen. |
| El líquido prostático confiere al semen un aspecto _________ y el líquido de las vesículas seminales y de las glándulas mucosas, la consistencia _____________. | El líquido prostático confiere al semen un aspecto lechoso y el líquido de las vesículas seminales y de las glándulas mucosas, la consistencia mucoide. |
| También, una proteína coagulante del líquido prostático hace que el fibrinógeno del líquido de la vesícula seminal forme un débil coágulo de fibrina que mantiene el semen en las regiones profundas de la vagina, donde está situado el cuello uterino. | El coágulo se disuelve durante los 15 a 30 min siguientes, debido a la lisis por la fibrinolisina formada a partir de la profibrinolisina prostática. En los primeros minutos siguientes a la eyaculación, los espermatozoides permanecen relativamente inmóviles, lo que podría deberse a la viscosidad del coágulo. A medida que este se disuelve, los espermatozoides adquieren una gran movilidad. |
| Aunque los espermatozoides pueden sobrevivir muchas semanas en los conductos genitales masculinos, una vez eyaculados en el semen su supervivencia máxima es solo de | 24 a 48 h a la temperatura corporal. Sin embargo, a bajas temperaturas puede almacenarse semen durante varias semanas y se han conservado espermatozoides durante años conservados a temperaturas inferiores a –100 °C. |
| *La «capacitación» de los espermatozoides es necesaria para | la fecundación del óvulo* |
| Aunque se dice que los espermatozoides están «maduros» cuando abandonan el epidídimo, su actividad permanece controlada por múltiples factores inhibidores secretados por los epitelios de los conductos genitales. Por tanto, inmediatamente después de su expulsión en el semen, son incapaces de fecundar el óvulo. Sin embargo, al entrar en contacto con los líquidos del aparato genital femenino, se producen múltiples cambios que activan a los espermatozoides para los procesos finales de la fecundación. Este conjunto de cambios recibe el nombre de | capacitación de los espermatozoides y suele tardar de 1 a 10 h en producirse. |
| Algunas de las modificaciones que se cree tienen lugar durante la capacitación de los espermatozoides son: | 1. Los líquidos del útero y de las trompas de Falopio eliminan los diversos factores inhibidores que mantenían reprimida la actividad de los espermatozoides en los conductos genitales masculinos. 2. Mientras los espermatozoides permanecen en el líquido de los conductos genitales masculinos están expuestos a numerosas vesículas flotantes de los túbulos seminíferos que contienen grandes cantidades de colesterol. Este colesterol se añade de manera continua a la membrana celular que reviste el acrosoma del espermatozoide, fortaleciéndola e impidiendo la liberación de sus enzimas. Tras la eyaculación, los espermatozoides depositados en la vagina nadan hacia arriba en el líquido uterino, alejándose de las vesículas de colesterol, y pierden poco a poco su exceso de colesterol en unas pocas horas. Al hacerlo, la membrana de la cabeza del espermatozoide (el acrosoma) se debilita mucho. |
| Algunas de las modificaciones que se cree tienen lugar durante la capacitación de los espermatozoides son: | 3. La membrana del espermatozoide se hace también mucho más permeable a los iones calcio, de forma que ahora penetra abundante calcio en el espermatozoide y modifica la actividad del flagelo, haciendo que adquiera una potente actividad de latigazo, en lugar del débil movimiento ondulante anterior. Además, es probable que los iones calcio produzcan alteraciones de la membrana celular que reviste la punta del acrosoma, facilitando la liberación de sus enzimas con rapidez y facilidad cuando el espermatozoide penetra en la masa de células de la granulosa que rodean al óvulo, e incluso más aún cuando trata de perforar la zona pelúcida del óvulo. |
| Enzimas del acrosoma, la «reacción del acrosoma» y la penetración en el óvulo. Almacenadas en el acrosoma del espermatozoide hay grandes cantidades de | hialuronidasa y de enzimas proteolíticas. |
| La hialuronidasa despolimeriza los polímeros de ácido hialurónico del cemento intercelular que mantiene unidas a las células de la granulosa del ovario. | Las enzimas proteolíticas digieren las proteínas de los elementos estructurales de los tejidos que todavía permanecen adheridos al óvulo. |
| Cuando el óvulo es expulsado del folículo ovárico hacia la trompa de Falopio, lleva consigo muchas capas de células de la granulosa. Antes de que un espermatozoide pueda fecundarlo, deberá disolver esta capa de células de la granulosa y después deberá penetrar a través de la densa cubierta del propio óvulo, la zona pelúcida. | Para conseguir esta penetración, comienzan a liberarse pequeñas cantidades de las enzimas del acrosoma. Se cree que una de ellas, la hialuronidasa, es especialmente importante para abrir camino entre las células de la granulosa de forma que el espermatozoide pueda alcanzar el óvulo. |
| Al llegar a la zona pelúcida del óvulo, la membrana anterior del espermatozoide se une de forma específica a proteínas receptoras de la zona pelúcida. Después, todo el acrosoma se disuelve con rapidez y se liberan de inmediato todas las enzimas. En cuestión de minutos estas enzimas abren una vía de penetración para el paso de la cabeza del espermatozoide a través de la zona pelúcida hasta el interior del óvulo. | En otros 30 min se fusionan las membranas de la cabeza del espermatozoide y del ovocito, formando una sola célula. Al mismo tiempo, el material genético del espermatozoide y del ovocito se combina para formar un genoma completamente nuevo, que contiene un número igual de cromosomas y genes del padre y de la madre. Este es el proceso de fecundación o fertilización. |
| *¿Por qué solo penetra un espermatozoide en el ovocito?* Habiendo tantos espermatozoides, ¿por qué solo penetra uno en el ovocito? La razón no se conoce del todo, pero pocos minutos después de la penetración del primer espermatozoide en la zona pelúcida del óvulo, iones calcio difunden a través de la membrana del ovocito y hacen que este libere por exocitosis numerosos gránulos corticales al espacio perivitelino. | Estos gránulos contienen sustancias que impregnan todas las porciones de la zona pelúcida e impiden la fijación de nuevos espermatozoides, e incluso hacen que se desprendan aquellos que ya se han unido. En cualquier caso, casi nunca penetra más de un espermatozoide en el ovocito durante la fecundación. |
| *Espermatogenia anormal y fertilidad masculina* El epitelio de los túbulos seminíferos puede destruirse por varias enfermedades. Por ejemplo. | La orquitis (inflamación) bilateral provocada por la parotiditis causa esterilidad en algunos hombres afectados. Asimismo, muchos niños varones nacen con una degeneración del epitelio tubular secundaria a la estenosis de los conductos genitales o de otras anomalías. Por último, otra causa de esterilidad, que suele ser transitoria, es la temperatura excesiva de los testículos. |
| Efecto de la temperatura sobre la espermatogenia | El aumento de la temperatura de los testículos puede impedir la espermatogenia y causar la degeneración de la mayor parte de las células de los túbulos seminíferos, además de las espermatogonias. Se ha afirmado repetidas veces que los testículos están situados en el escroto colgante para que puedan mantener una temperatura inferior a la temperatura interna del cuerpo, aunque habitualmente solo unos 2 °C menos. En los días fríos, los reflejos escrotales hacen que la musculatura del escroto se contraiga, acercando los testículos al cuerpo para mantener esta diferencia de 2 °C. Por tanto, el escroto actúa como un mecanismo de enfriamiento de los testículos (pero un enfriamiento controlado), sin el cual la espermatogenia podría ser deficiente cuando el clima es muy caluroso. |
| Criptorquidia | Criptorquidia significa falta de descenso de un testículo desde el abdomen al escroto en el período perinatal. Durante el desarrollo del feto masculino, los testículos se forman a partir de las crestas genitales en el abdomen. Sin embargo, entre 3 semanas y 1 mes antes del nacimiento del niño, los testículos descienden a través de los conductos inguinales al escroto. A veces, este descenso no se produce o es incompleto, de forma que uno o ambos testículos permanecen en el abdomen, en el conducto inguinal o en otro punto de la ruta de descenso. |
| Un testículo que permanece en el interior de la cavidad abdominal es incapaz de formar espermatozoides. El epitelio tubular degenera, dejando solo las estructuras intersticiales del órgano. Se ha afirmado que los pocos grados más de temperatura que existen en el abdomen respecto al escroto bastan para causar la degeneración del epitelio tubular y, en consecuencia, provocar esterilidad, pero este efecto no es totalmente seguro. | Sin embargo, por esta razón, pueden realizarse operaciones para recolocar los testículos criptorquídicos desde la cavidad abdominal al interior del escroto antes del inicio de la vida sexual adulta en niños con testículos no descendidos. La secreción de testosterona por los testículos fetales es el estímulo normal que provoca el descenso de los testículos al escroto desde el abdomen. Por ello, muchos, si no la mayoría, de los casos de criptorquidia se deben a testículos anormales que no son capaces de secretar la testosterona suficiente. En los pacientes con esta forma de criptorquidia es improbable que la cirugía tenga éxito. |
| *Efecto del recuento de espermatozoides sobre la fertilidad* | La cantidad de semen eyaculado en cada coito es, como promedio, de 3,5 ml y en cada mililitro de semen hay un promedio de unos 120 millones de espermatozoides, aunque incluso en varones «normales» el recuento puede variar entre 35 y 200 millones. Esto significa que en los mililitros de cada eyaculación hay un promedio de 400 millones de espermatozoides. Cuando el número de espermatozoides por mililitro cae por debajo de unos 20 millones, es probable que la persona no sea fértil. Por tanto, a pesar de que solo se necesita un espermatozoide para fecundar al óvulo, por razones que no se conocen por completo, el eyaculado debe contener un ingente número de ellos para que uno solo fecunde al óvulo. |
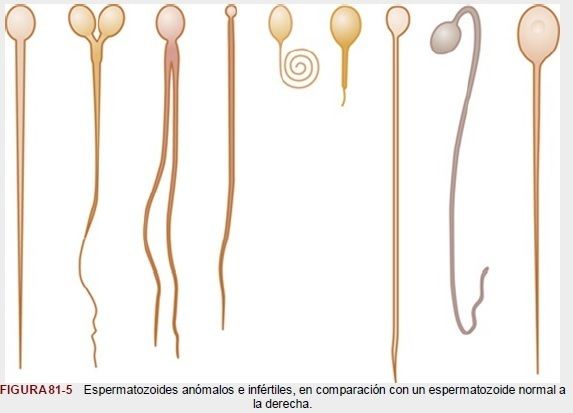
| *Efecto de la morfología y la motilidad de los espermatozoides sobre la fertilidad* A veces, un varón con un recuento normal de espermatozoides es estéril. Cuando se produce esta situación, puede encontrarse que hasta la mitad de los espermatozoides presentan anomalías morfológicas, con dos cabezas, con cabezas de forma anormal o colas anormales, como muestra la figura 81-5. En otros casos, la estructura de los espermatozoides es normal pero, por razones no conocidas, son completa o relativamente inmóviles. Siempre que la forma de la mayoría de los espermatozoides sea anormal o no puedan moverse, será probable que el varón sea estéril, aunque el resto de los espermatozoides tengan un aspecto normal. |
Image:
Esp (binary/octet-stream)
|
| *Acto sexual masculino* Estímulo neuronal para el rendimiento del acto sexual masculino La fuente más importante de señales nerviosas sensitivas para la iniciación del acto sexual masculino es el | glande del pene. |
| El glande contiene un órgano sensitivo muy sensible que transmite al sistema nervioso central una modalidad especial de sensación denominada | sensación sexual. La acción de masaje del glande en la relación sexual estimula los órganos sensitivos terminales y las señales sexuales, a su vez, se propagan a través del nervio pudendo y después, por el plexo sacro, a la porción sacra de la médula espinal y por último ascienden a través de la médula hasta proyectarse en áreas no definidas del encéfalo. |
| Los impulsos también pueden penetrar en la médula espinal procedentes de áreas próximas al pene para ayudar a estimular el acto sexual. Por ejemplo, la estimulación del epitelio anal, el escroto y las estructuras perineales en general puede enviar señales a la médula que contribuyen a la sensación sexual. Las sensaciones sexuales pueden originarse incluso en estructuras internas, como en zonas de la uretra, la vejiga, la próstata, las vesículas seminales, los testículos y el conducto deferente. De hecho, una de las causas del «impulso sexual» es que los órganos sexuales estén llenos de secreciones. | La infección y la inflamación leves de estos órganos sexuales pueden provocar en ocasiones un deseo sexual casi ininterrumpido y algunos «afrodisíacos», como las cantáridas, irritan las mucosas vesical y uretral lo que induce inflamación y congestión vascular. |
| *Elemento psíquico de la estimulación sexual masculina* | Los estímulos psicológicos adecuados pueden facilitar mucho la capacidad de una persona para realizar el acto sexual. Los simples pensamientos de contenido sexual o incluso el hecho de soñar que se está realizando el coito pueden hacer que se produzca el acto sexual masculino, culminando en la eyaculación. De hecho, en muchos varones se producen eyaculaciones nocturnas, llamadas «sueños húmedos», en algunas etapas de la vida sexual, especialmente durante la adolescencia. |
| *Integración del acto sexual masculino en la médula espinal* Aunque los factores psicológicos desempeñan habitualmente un papel importante en el acto sexual masculino y pueden iniciarlo o inhibirlo, es probable que la función encefálica no sea necesaria para su rendimiento, debido a que la estimulación genital adecuada puede causar, tras la sección de la médula espinal por encima de la región lumbar, la eyaculación en algunos animales y a veces en el ser humano. | Por tanto, el acto sexual masculino es el resultado de mecanismos reflejos intrínsecos integrados en la médula espinal sacra y lumbar, y estos mecanismos pueden iniciarse tanto por estimulación psicológica del encéfalo como por una estimulación sexual real de los órganos sexuales, aunque lo habitual es que ocurra debido a una combinación de ambas. |
| Etapas del acto sexual masculino | Erección: función de los nervios parasimpáticos La lubricación es una función parasimpática La emisión y la eyaculación son funciones de los nervios simpáticos |
| *Erección: función de los nervios parasimpáticos* La erección del pene es el primer efecto de la estimulación sexual masculina y el grado de erección es proporcional al grado de estimulación, sea psíquica o física. La erección se debe a los impulsos parasimpáticos que alcanzan el pene desde la porción sacra de la médula espinal a través de los nervios pélvicos. Se cree que, a diferencia de la mayoría de fibras parasimpáticas, estas secretan | óxido nítrico, péptido intestinal vasoactivo o ambos, además de acetilcolina. |
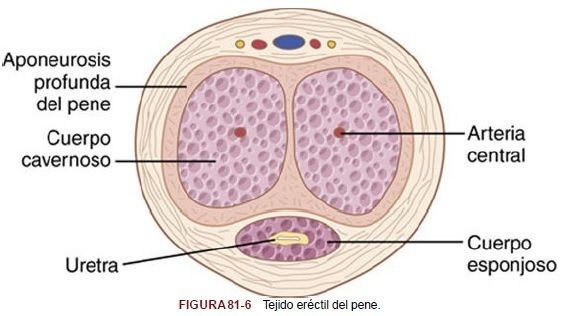
| El óxido nítrico activa la enzima guanililo ciclasa, lo que provoca el aumento de la formación de monofosfato de guanosina cíclico (GMPc). | El GMPc relaja especialmente las arterias del pene, así como la red trabecular de fibras musculares lisas del tejido eréctil de los cuerpos cavernosos y del cuerpo esponjoso del cuerpo del pene, que se muestran en la figura 81-6. Cuando los músculos lisos vasculares se relajan, el flujo sanguíneo en el pene aumenta, lo que provoca la liberación de óxido nítrico desde las células endoteliales vasculares y ulterior vasodilatación. |
|
Image:
Aaa (binary/octet-stream)
|
El tejido eréctil del pene no es otra cosa que un conjunto de grandes sinusoides cavernosos, que en condiciones normales contienen poca sangre, pero que experimentan una gran dilatación cuando la sangre arterial fluye a su interior a presión mientras el flujo venoso está parcialmente ocluido. Además, los cuerpos eréctiles, en especial los dos cuerpos cavernosos, están también rodeados de fuertes revestimientos fibrosos; por tanto, la elevada presión en el interior de los sinusoides provoca un abombamiento del tejido eréctil, de forma tal que el pene se endurece y se alarga, un fenómeno que se denomina erección. |
| *La lubricación es una función parasimpática* Durante la estimulación sexual, los impulsos parasimpáticos, además de promover la erección, hacen que las glándulas uretrales y bulbouretrales secreten moco. Este moco fluye a través de la uretra durante la cópula y ayuda a la lubricación del coito. | No obstante, la mayor parte de dicha lubricación procede de los órganos sexuales femeninos más que de los masculinos. Sin una lubricación satisfactoria, el acto sexual masculino rara vez tiene éxito, debido a que el coito sin lubricación provoca sensaciones de raspado, dolorosas, que inhiben en lugar de excitar las sensaciones sexuales. |
| *La emisión y la eyaculación son funciones de los nervios simpáticos* La emisión y la eyaculación son la culminación del acto sexual masculino. | Cuando el estímulo sexual es extremadamente intenso, los centros reflejos de la médula espinal comienzan a emitir impulsos simpáticos que abandonan la médula al nivel de T12 a L2 y pasan a los órganos genitales por los plexos nerviosos simpáticos hipogástricos y pélvicos para iniciar la emisión, el preludio de la eyaculación. |
| La emisión comienza con | la contracción del conducto deferente y de la ampolla para provocar la expulsión de los espermatozoides a la uretra interna.Después, las contracciones del revestimiento muscular de la glándula prostática, seguidas de la contracción de las vesículas seminales, expelen el líquido prostático y seminal hacia la uretra, empujando hacia adelante a los espermatozoides. Todos estos líquidos se mezclan en la uretra interna con el moco ya secretado por las glándulas bulbouretrales para formar el semen. El proceso, hasta este punto, es la emisión. |
| El llenado de la uretra interna por el semen desencadena señales sensitivas que se transmiten a través de los nervios pudendos a las zonas sacras de la médula, produciendo una sensación de repentina repleción de los órganos genitales internos. Estas señales sensitivas estimulan también la contracción rítmica de los órganos genitales internos y causan la contracción de los músculos isquiocavernosos y bulbocavernosos que comprimen las bases del tejido eréctil peniano. | La conjunción de todos estos efectos unidos determina un aumento rítmico, en oleadas, de la presión en el tejido eréctil del pene, en los conductos genitales y en la uretra, que «eyaculan» el semen desde la uretra al exterior. Este proceso final se denomina eyaculación. Al mismo tiempo, las contracciones rítmicas de los músculos pélvicos e incluso de algunos músculos del tronco producen movimientos de vaivén de la pelvis y del pene, que ayudan también a propulsar el semen a las zonas más profundas de la vagina e incluso ligeramente al interior del cuello uterino. |
| Este período completo de la emisión y eyaculación se denomina | orgasmo masculino. Al terminar, la excitación sexual del varón desaparece casi por completo en 1 a 2 min y la erección termina, un proceso denominado resolución. |
| *Testosterona y otras hormonas masculinas Secreción, metabolismo y química de las hormonas masculinas Secreción de testosterona por las células intersticiales de Leydig de los testículos* Los testículos secretan varias hormonas sexuales masculinas, que en conjunto reciben el nombre de andrógenos y que son | la testosterona, la dihidrotestosterona y la androstenodiona. |
| La cantidad de testosterona es tan superior a la de las demás que se puede considerar la hormona testicular más importante, si bien buena parte de la testosterona se convierte en los tejidos efectores en | dihidrotestosterona, una hormona más activa. |
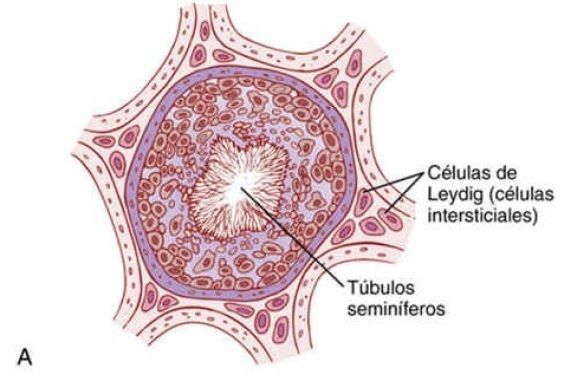
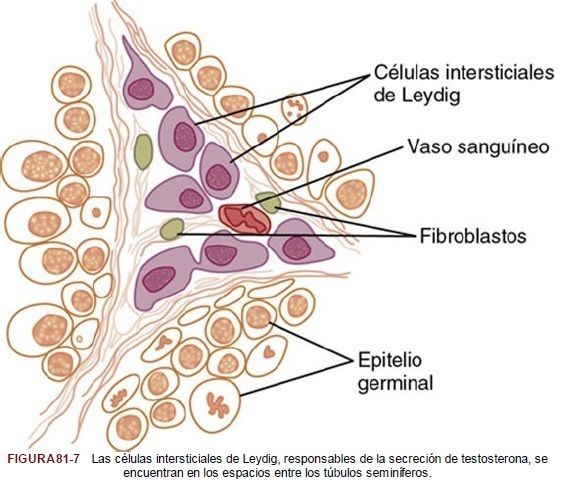
| La testosterona se produce en las |
células intersticiales de Leydig, que están situadas en los intersticios existentes entre los túbulos seminíferos y que constituyen alrededor del 20% de la masa del testículo adulto, como refleja la figura 81-7.
Image:
Cv (binary/octet-stream)
|
| Las células de Leydig son casi inexistentes en los testículos durante la niñez, en la que los testículos apenas secretan testosterona, pero son muy numerosas en el recién nacido varón durante los primeros meses de vida y en el varón adulto en cualquier momento después de la pubertad; en estas dos etapas de la vida, los testículos secretan grandes cantidades de testosterona. | Además, cuando se desarrollan tumores de las células intersticiales de Leydig, se producen grandes cantidades de testosterona. Por último, cuando el epitelio germinativo de los testículos se destruye a causa de la radioterapia o de un calor excesivo, las células de Leydig, que son más resistentes, continúan fabricando testosterona. |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Want to create your own Flashcards for free with GoConqr? Learn more.