12682231
Description
Mind Map by Rosa Isela Angeles López, updated more than 1 year ago
|
|
Created by Rosa Isela Angeles López
almost 7 years ago
|
|
FIJACIÓN DEL CO2 EN LAS PLANTAS C3, C4 Y CAM
- PLANTAS
C3
- Casi el 85% de las especies de plantas
son C3, se dice que son plantas que no
tienen adaptaciones fotosintéticas
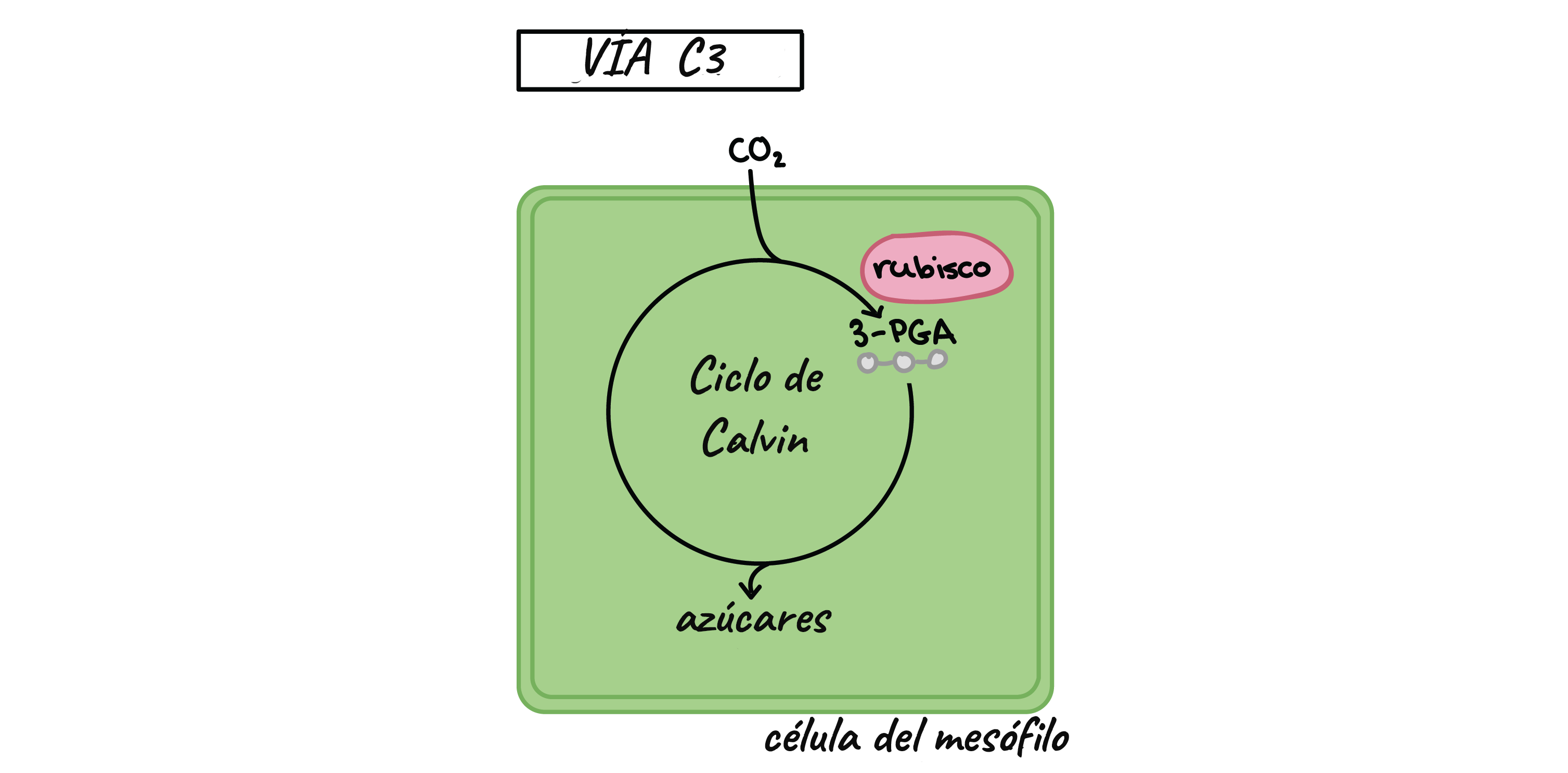
- El primer compuesto
orgánico que incorpora el
Co2 es 3 fosfoglicerato
- Interviene el ciclo de Calvin-Benson. En
este ciclo se distinguen LAS SIGUIENTES
ETAPAS
- Fase de Carboxilación
- Esta es una fase enzimática que consiste de una
reacción mediante la cual el CO2 es adicionado
a un azúcar de 5 carbonos, la ribosa 1,5
bi-fosfato (RuBP) para formar dos moléculas de
ácido fosfoglicérico (PGA) de tres carbonos.
- Esta reacción es catalizada por la enzima
ribosa 1,5 bifosfato carboxilasa/oxigenasa
(Rubisco)
- Esta reacción es catalizada por la enzima
ribosa 1,5 bifosfato carboxilasa/oxigenasa
(Rubisco)
- Esta es una fase enzimática que consiste de una
reacción mediante la cual el CO2 es adicionado
a un azúcar de 5 carbonos, la ribosa 1,5
bi-fosfato (RuBP) para formar dos moléculas de
ácido fosfoglicérico (PGA) de tres carbonos.
- Fase de Reducción
- En esta fase, el PGA (ácido orgánico)
formado por la adición de CO2 a la
ribosa 1,5 bifosfato es convertido
(reducido) en un azúcar de 3 carbonos
(Triosa-P). En este proceso es necesario
utilizar la energía del “poder reductor”
del NADPH2 e el ATP.
- La reacción se da en dos etapas, la primera de fosforilación,
adicionando un P del ATP y a continuación reduciendo como
NADPH2. El poder reductor del NADPH2 es utilizado para
transformar el grupo ácido del PGA en el grupo aldehído de la
triosa-P; el ATP es necesario para suplir energía extra a fin de
ejecutar esta etapa.
- Desde el momento que el CO2 ha sido reducido al nivel del azúcar de 3
carbonos (triosa-P), la parte que conserva la energía de la fotosíntesis fue
ejecutada. Luego de eso, es necesario regenerar la molécula inicial aceptora
de CO2 , esto es, la ribosa 1,5 bifosfato, a fin de que la fijación del CO2
continúe indefinidamente (fase de regeneración) y transformar la triosa-P en
azúcares más complejos, carbohidratos, grasas, aminoácidos, etc. (fase de
síntesis de productos).
- Desde el momento que el CO2 ha sido reducido al nivel del azúcar de 3
carbonos (triosa-P), la parte que conserva la energía de la fotosíntesis fue
ejecutada. Luego de eso, es necesario regenerar la molécula inicial aceptora
de CO2 , esto es, la ribosa 1,5 bifosfato, a fin de que la fijación del CO2
continúe indefinidamente (fase de regeneración) y transformar la triosa-P en
azúcares más complejos, carbohidratos, grasas, aminoácidos, etc. (fase de
síntesis de productos).
- La reacción se da en dos etapas, la primera de fosforilación,
adicionando un P del ATP y a continuación reduciendo como
NADPH2. El poder reductor del NADPH2 es utilizado para
transformar el grupo ácido del PGA en el grupo aldehído de la
triosa-P; el ATP es necesario para suplir energía extra a fin de
ejecutar esta etapa.
- En esta fase, el PGA (ácido orgánico)
formado por la adición de CO2 a la
ribosa 1,5 bifosfato es convertido
(reducido) en un azúcar de 3 carbonos
(Triosa-P). En este proceso es necesario
utilizar la energía del “poder reductor”
del NADPH2 e el ATP.
- Fase de Regeneración
- El aceptor inicial de CO2, RuBP es
regenerado para posteriores reacciones de
fijación, a través de una serie compleja de
reacciones involucrando azúcares
fosfatados con 3, 4, 5, 6 y 7 carbonos.
- El aceptor inicial de CO2, RuBP es
regenerado para posteriores reacciones de
fijación, a través de una serie compleja de
reacciones involucrando azúcares
fosfatados con 3, 4, 5, 6 y 7 carbonos.
- Fase de Síntesis de Productos
- Los productos finales de la fotosíntesis son
considerados primariamente como azúcares y
otros carbohidratos, más grasas, ácidos grasos,
aminoácidos y ácidos orgánicos han sido
también adminitos como sintetizados en la
fijación fotosintética del carbono.
- Los productos finales de la fotosíntesis son
considerados primariamente como azúcares y
otros carbohidratos, más grasas, ácidos grasos,
aminoácidos y ácidos orgánicos han sido
también adminitos como sintetizados en la
fijación fotosintética del carbono.
- Fase de Carboxilación
- El primer compuesto
orgánico que incorpora el
Co2 es 3 fosfoglicerato
- Casi el 85% de las especies de plantas
son C3, se dice que son plantas que no
tienen adaptaciones fotosintéticas
- PLANTAS POR LA RUTA
C4, LA VÍA DE 4
CARBONOS O LA VÍA DE
HATCH-SLACK
- Cerca del 3% de todas
plantas vasculares este
sistemas fotosinteticos,
son comunes en los
hábitats cálidos y menos
abundantes en zonas
frescas
- Se realiza mediante
las reacciones del ciclo
de Calvin- Benson.
- Su primer producto formado es
OXALACETATO (4C)
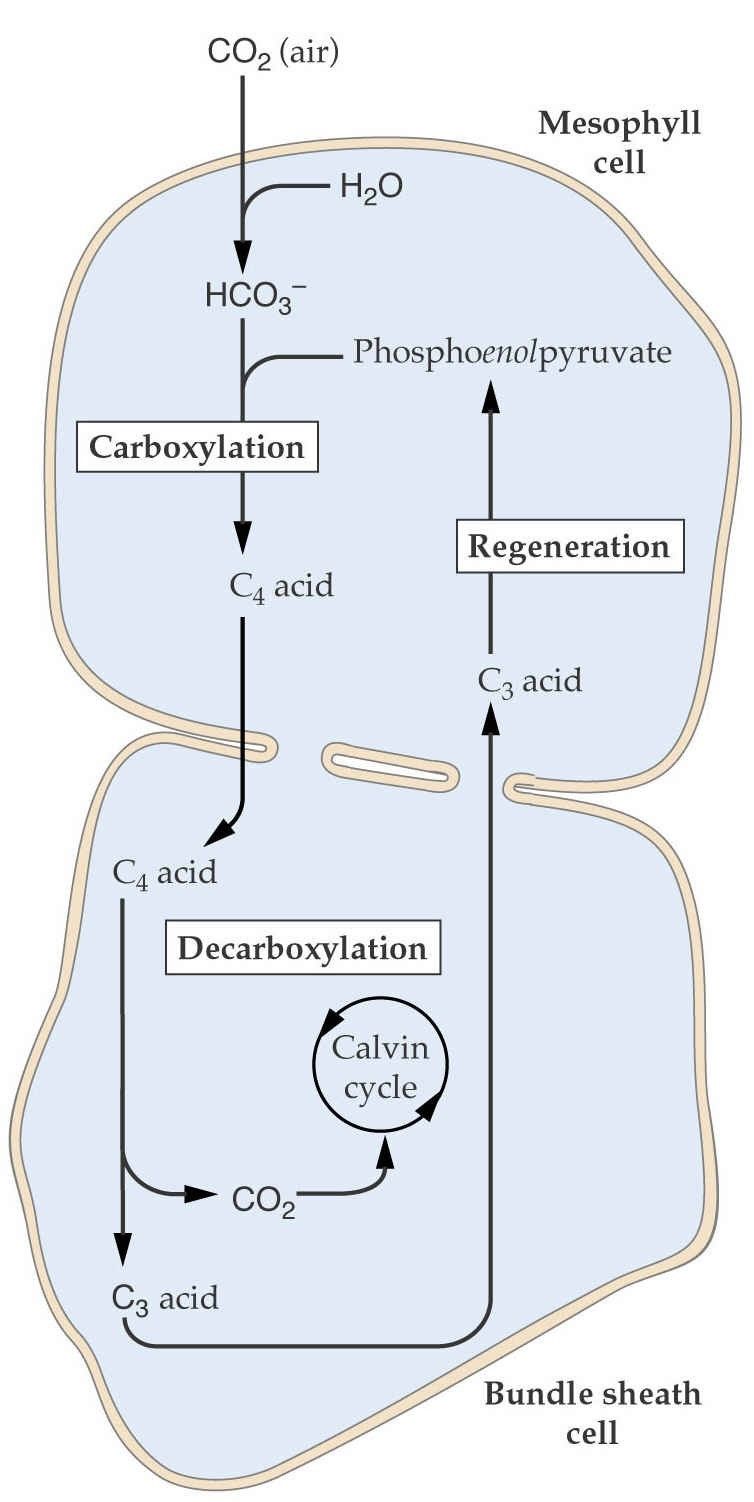
- En las plantas C4, la fijación inicial del CO2
ocurre en las células mesofílicas. En el citosol de
estas células, el CO2 reacciona con el
fosfoenolpiruvato vía enzima fosfoenolpiruvato
carboxilasa (PEP carboxilasa) para formar
oxalacetato. Existe elevada concentración de
PEP carboxilasa en las células mesofílicas.
Subsecuentemente, el oxalacetato puede ser
reducido a malato con utilización del NADPH2 o
puede ser aminado en aspartato.
- Asimilación de CO2,
- en células del mesófilo,
que se transforma
rápidamente mediante un
proceso de hidratación en
HCO3¯ el cual se fija al
fosfoenolpiruvato en
una reacción de
carboxilación catalizada
por la fosfoenolpiruvato
carboxilasa para dar
oxalacetato.
- en células del mesófilo,
que se transforma
rápidamente mediante un
proceso de hidratación en
HCO3¯ el cual se fija al
fosfoenolpiruvato en
una reacción de
carboxilación catalizada
por la fosfoenolpiruvato
carboxilasa para dar
oxalacetato.
- Transporte
- Transporte de los ácidos C-4
(málico o aspártico) a las células
de la vaina.
- Transporte de los ácidos C-4
(málico o aspártico) a las células
de la vaina.
- Decarboxilación
- Decarboxilación de los ácidos C-4 en las
células de la vaina y generación de CO2
que entra al ciclo de Calvin.
- Decarboxilación de los ácidos C-4 en las
células de la vaina y generación de CO2
que entra al ciclo de Calvin.
- Regreso a las células del mesófilo de los ácidos
C3 (piruvato o alanina) y posterior
regeneración del aceptor inicial,
fosfoenolpiruvato
- Asimilación de CO2,
- En las plantas C4, la fijación inicial del CO2
ocurre en las células mesofílicas. En el citosol de
estas células, el CO2 reacciona con el
fosfoenolpiruvato vía enzima fosfoenolpiruvato
carboxilasa (PEP carboxilasa) para formar
oxalacetato. Existe elevada concentración de
PEP carboxilasa en las células mesofílicas.
Subsecuentemente, el oxalacetato puede ser
reducido a malato con utilización del NADPH2 o
puede ser aminado en aspartato.
- Su primer producto formado es
OXALACETATO (4C)
- Algunas especies de plantas y muchas gramíneas de
regiones tropicales (maíz, sorgo, caña de azúcar), son
capaces de fijar CO2 en compuestos de 4 carbonos,
como oxalacetato, malato y aspartato, además de la
reducción operada por el ciclo C3 de Calvin
- Se realiza mediante
las reacciones del ciclo
de Calvin- Benson.
- Cerca del 3% de todas
plantas vasculares este
sistemas fotosinteticos,
son comunes en los
hábitats cálidos y menos
abundantes en zonas
frescas
- PLANTAS CON
METABOLISMO ÁCIDO DE
LAS CRASULÁCEAS
- Predominan en zonas
muy cálidas y secas como
los desiertos.
- Dividen la fotosintésis de dos
tiempos ya que en el día
cierran los estromas y
durante la noche abren los
estromas
- El primer producto
formado es el
Oxalacetato C4
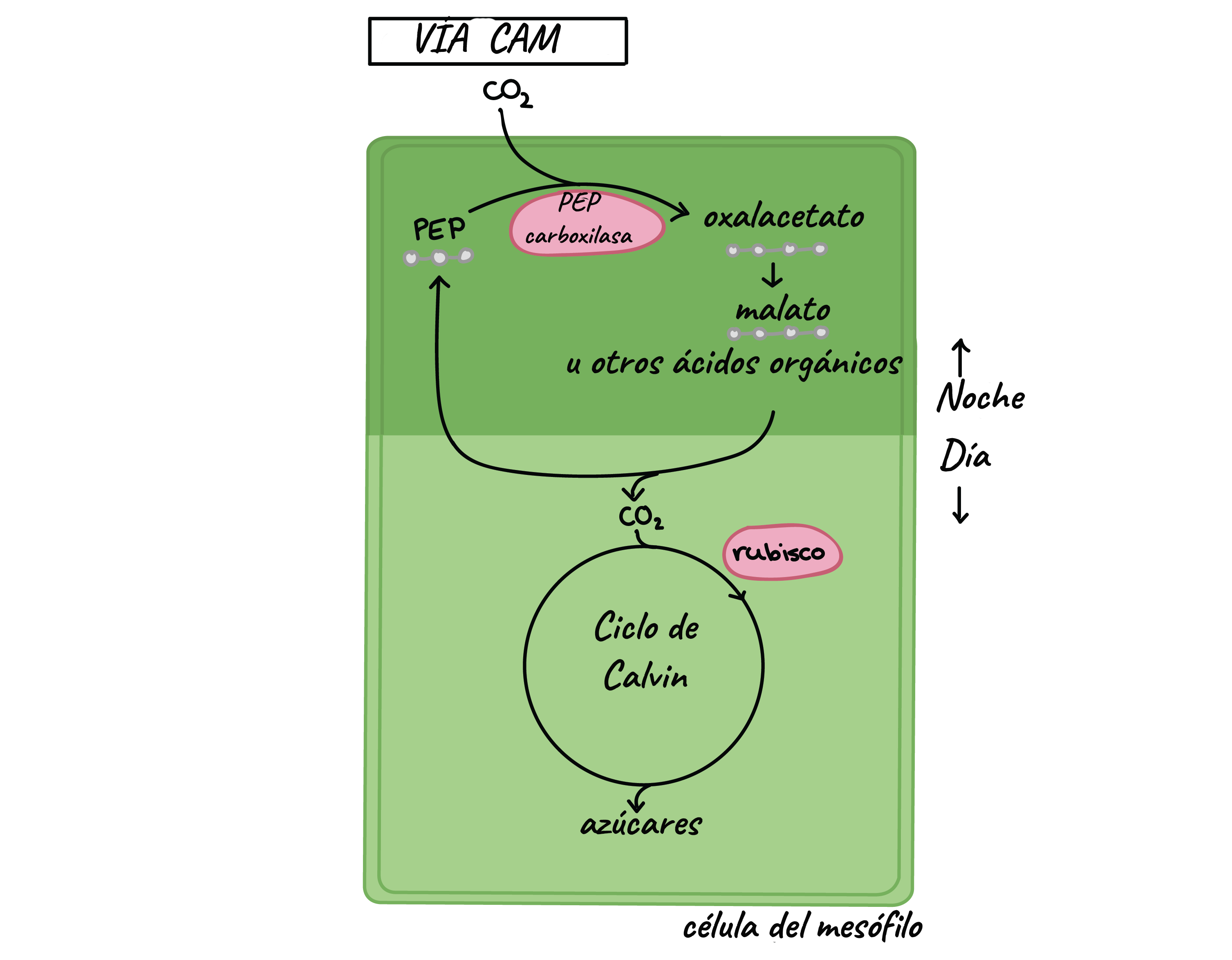
- El mecanismo de fijación del CO2 en las
plantas CAM y en muchos aspectos similar
al mecanismo de fijación de las plantas
C4. Las plantas CAM, presentan dos vías
de fijación del CO2, una fijación inicial
por la PEP carboxilasa y luego de una
re-fijación vía Rubisco. Sin embargo, en
las CAM las dos vías de fijación de CO2
están separadas temporalmente.
- Inicialmente el CO2 es fijado por la
noche vía enzima PEP-carboxilasa,
utilizando PEP como aceptor y
formando oxalacetato que en
seguida es reducido a malato. El
malato se acumula en el vacuolo.
La acumulación de malato durante
la noche, equivalente al CO2 fijado,
provoca la acidificación nocturna
de la hoja.
- Al día siguiente, con los estomas cerrados,
el malato sale del vacuolo y se
descarboxila por acción de la NAPD
enzima málica, en piruvato y CO2. El CO2
liberado internamente no escapa de la
hoja y es refijado vía rubisco (ciclo de
calvin). La elevada concentración interna
de CO2 que se genera favorece la
actividad caboxilativa de la Rubisco y
reprime la oxigenación fotorespiratoria de
la RuBP
- Inicialmente el CO2 es fijado por la
noche vía enzima PEP-carboxilasa,
utilizando PEP como aceptor y
formando oxalacetato que en
seguida es reducido a malato. El
malato se acumula en el vacuolo.
La acumulación de malato durante
la noche, equivalente al CO2 fijado,
provoca la acidificación nocturna
de la hoja.
- El mecanismo de fijación del CO2 en las
plantas CAM y en muchos aspectos similar
al mecanismo de fijación de las plantas
C4. Las plantas CAM, presentan dos vías
de fijación del CO2, una fijación inicial
por la PEP carboxilasa y luego de una
re-fijación vía Rubisco. Sin embargo, en
las CAM las dos vías de fijación de CO2
están separadas temporalmente.
- El primer producto
formado es el
Oxalacetato C4
- Dividen la fotosintésis de dos
tiempos ya que en el día
cierran los estromas y
durante la noche abren los
estromas
- Predominan en zonas
muy cálidas y secas como
los desiertos.
Media attachments
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Want to create your own Mind Maps for free with GoConqr? Learn more.