
13788271
Description
Biología Celular y
Molecular II
- MECANISMOS DE COMUNICACIÓN
CELULAR
- PRINCIPIOS GENERALES DE LA
COMUNICACIÓN CELULAR
- hace mucho antes que aparecieran los organismos
pluricelulares los organismos unicelulares ya habian
desarrollado un mecanismo que les permitia
repsonder a cambios químicos o físicos de su entorno
y responder a la presencia de otras células
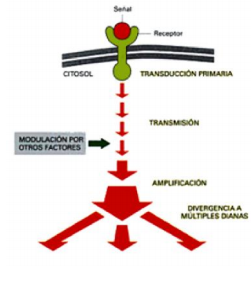
- CASCADA DE
SEÑALIZACIÓN
- Secuencia de reacciones de proteínas que actúan en cadena
para transmitir señal en la célula. Las reacciones incluyen
frecuentemente fosforilaciones y desfosforilaciones.
- Secuencia de reacciones de proteínas que actúan en cadena
para transmitir señal en la célula. Las reacciones incluyen
frecuentemente fosforilaciones y desfosforilaciones.
- hace mucho antes que aparecieran los organismos
pluricelulares los organismos unicelulares ya habian
desarrollado un mecanismo que les permitia
repsonder a cambios químicos o físicos de su entorno
y responder a la presencia de otras células
- LAS MOLÉCULAS SEÑAL EXTRACELULARES
SE UNEN A RECEPTORES ESPECÍFICOS
- Los organismos pluricelulares se
comunican mediante diferentes
tipos de moleculas señal como
- Proteínas
- retinoides
- aminoádos
- nucleótidos
- esteroides
- pueden ser liberadas por las células señalizadoras y otras por difusión.
Las proteínas transmembrana pueden actuar como señalizadoras de
esta forma pueden ser liberados a la superficie celular mediante rotura
proteolítica y así, actuar a la distancia
- Molécula señal,
primer
mensajero o
ligando
- •Función de activar al receptor en las
células blanco. •Luego, las células
blanco modifican o degradan al ligando
dando término a la respuesta.
- Segundos
mensajeros:
- son mediadores
intracelulares, moléculas
señalizadoras de pequeño
tamaño.
- son mediadores
intracelulares, moléculas
señalizadoras de pequeño
tamaño.
- •Función de activar al receptor en las
células blanco. •Luego, las células
blanco modifican o degradan al ligando
dando término a la respuesta.
- Molécula señal,
primer
mensajero o
ligando
- Proteínas
- Los organismos pluricelulares se
comunican mediante diferentes
tipos de moleculas señal como
- FORMAS DE SEÑALIZACIÓN
INTERCELULAR
- a) Dependiente
de contacto
- b) Paracrina
- c) Sináptica
- d) Endocrina
- e) Autocrina
- a) Dependiente
de contacto
- RECEPTORES DE SUPERFICIE
CELULAR
- • Receptores acoplados a canales
iónicos
- • Receptores acoplados a proteínas
G:
- SEÑALIZACIÓN A TRAVÉS DE
RECEPTORES ACOPLADOS A PROTEINAS G
- Segundos mensajeros de los
receptores acoplados a proteínas G
- SEÑALIZACIÓN A TRAVÉS DE
RECEPTORES ACOPLADOS A PROTEINAS G
- • Receptores acoplados a
enzimas
- • Receptores acoplados a canales
iónicos
- . La función principal de la comunicación celular
es la de adaptarse a los cambios que existen en
el medio que les rodea para sobrevivir a esos
cambios, gracias al fenómeno de la homeostasis
- Señalización a través de receptores de
superficie celular acoplados a proteínas G Y
mediadores intracelulares pequeños
- Los GPCR a través de proteínas G
activan o inactivan enzimas unidas a la
membrana plasmática o canales iónicos.
- Un incremento de los niveles de
AMP cíclico o de Ca afecta la célula
estimulando la proteína quinasa
- Un incremento de los niveles de
AMP cíclico o de Ca afecta la célula
estimulando la proteína quinasa
- La PKC,PKA y LAS CaM-
quinasas fosforilan proteínas
diana determinadas en residuos
serina y treonina alterando la
actividad de proteínas.
- Cada tipo celular tiene su propio
conjunto característico de proteínas
diana que son reguladas
- Cada tipo celular tiene su propio
conjunto característico de proteínas
diana que son reguladas
- Las diferentes respuestas
mediadas por GPCR son
desactivadas cuando el ligando
señal extracelular es eliminado.
- Los propios GPCR son
fosforilados por GRK lo que
hace que la arrestina se una.
- Los propios GPCR son
fosforilados por GRK lo que
hace que la arrestina se una.
- Los GPCR a través de proteínas G
activan o inactivan enzimas unidas a la
membrana plasmática o canales iónicos.
- Señalización a través
de receptores de
superficie celular
acoplados a enzimas
- Existen varias clase de receptores
incluyendo los receptores tirosina
quinasa RTK los receptores asociados a
tirosina quinasa los receptores a
serina/treonina quinasa y los receptores
asociados a histidina quinasa.
- La unión del ligando a RTK
induce a los receptores a la
fosforilacion cruzada en
varios residuos tirosina de
sus dominios citoplasmáticos.
- Receptores
intracelulares
- Son proteínas receptoras
que se encuentran al
interior de la célula.
- Son proteínas receptoras
que se encuentran al
interior de la célula.
- Receptores de
superficie
celular
- Son proteínas ancladas a
la membrana que se
unen al ligando en la
parte exterior de la célula
- Son proteínas ancladas a
la membrana que se
unen al ligando en la
parte exterior de la célula
- Receptores
intracelulares
- La unión del ligando a RTK
induce a los receptores a la
fosforilacion cruzada en
varios residuos tirosina de
sus dominios citoplasmáticos.
- Existen varias clase de receptores
incluyendo los receptores tirosina
quinasa RTK los receptores asociados a
tirosina quinasa los receptores a
serina/treonina quinasa y los receptores
asociados a histidina quinasa.
- Canales iónicos
activados por ligando
- Son canales de iones que abren en
respuesta a la unión de un ligando.
- Son canales de iones que abren en
respuesta a la unión de un ligando.
- Vías de señalización
dependientes de
proteólisis regulada
por proteínas latentes
reguladas de genes.
- La unión inicia una
vía de señalización
- Cuando un ligando se
une a un receptor de
superficie celular, el
dominio intracelular
del receptor cambia
de algún modo.
- Fosforilación
- Las proteínas pueden
activarse o desactivarse
de varias maneras.
- Las proteínas pueden
activarse o desactivarse
de varias maneras.
- Segundos mensajeros
- Moléculas no proteicas que
pasan la señal iniciada por
la unión de un ligando
- Iones de Ca
- AMP ciclico
- Inositol Fosfato
- Iones de Ca
- Moléculas no proteicas que
pasan la señal iniciada por
la unión de un ligando
- Fosforilación
- Cuando un ligando se
une a un receptor de
superficie celular, el
dominio intracelular
del receptor cambia
de algún modo.
- La unión inicia una
vía de señalización
- Señalización a través de receptores de
superficie celular acoplados a proteínas G Y
mediadores intracelulares pequeños
- PRINCIPIOS GENERALES DE LA
COMUNICACIÓN CELULAR
- CITOESQUELETO Y COMPORTAMIENTO CELULAR
- CILIOS
- son más largos que los cilios, una célula tiene
pocos o un solo flagelo.S on estructuras
móviles muy especializadas y eficientes
formadas por microtúbulos y dineína
- más cortos que los flagelos y están organizados
de forma parecida, pero su movimiento es
parecido al movimiento de una brazada.
- más cortos que los flagelos y están organizados
de forma parecida, pero su movimiento es
parecido al movimiento de una brazada.
- son más largos que los cilios, una célula tiene
pocos o un solo flagelo.S on estructuras
móviles muy especializadas y eficientes
formadas por microtúbulos y dineína
- FLAGELOS
- Ambas son estructuras apéndices
celulares filiformes que contienen un
haz de microtúbulos en su interior.
- se presentan en los espermatozoides
y en muchos protozoos, mediante su
movimiento ondulante permiten
nadar por los medios líquidos.
- se presentan en los espermatozoides
y en muchos protozoos, mediante su
movimiento ondulante permiten
nadar por los medios líquidos.
- Ambas son estructuras apéndices
celulares filiformes que contienen un
haz de microtúbulos en su interior.
- movimiento está
producido por el
batido de su parte
central llamado:
axonema
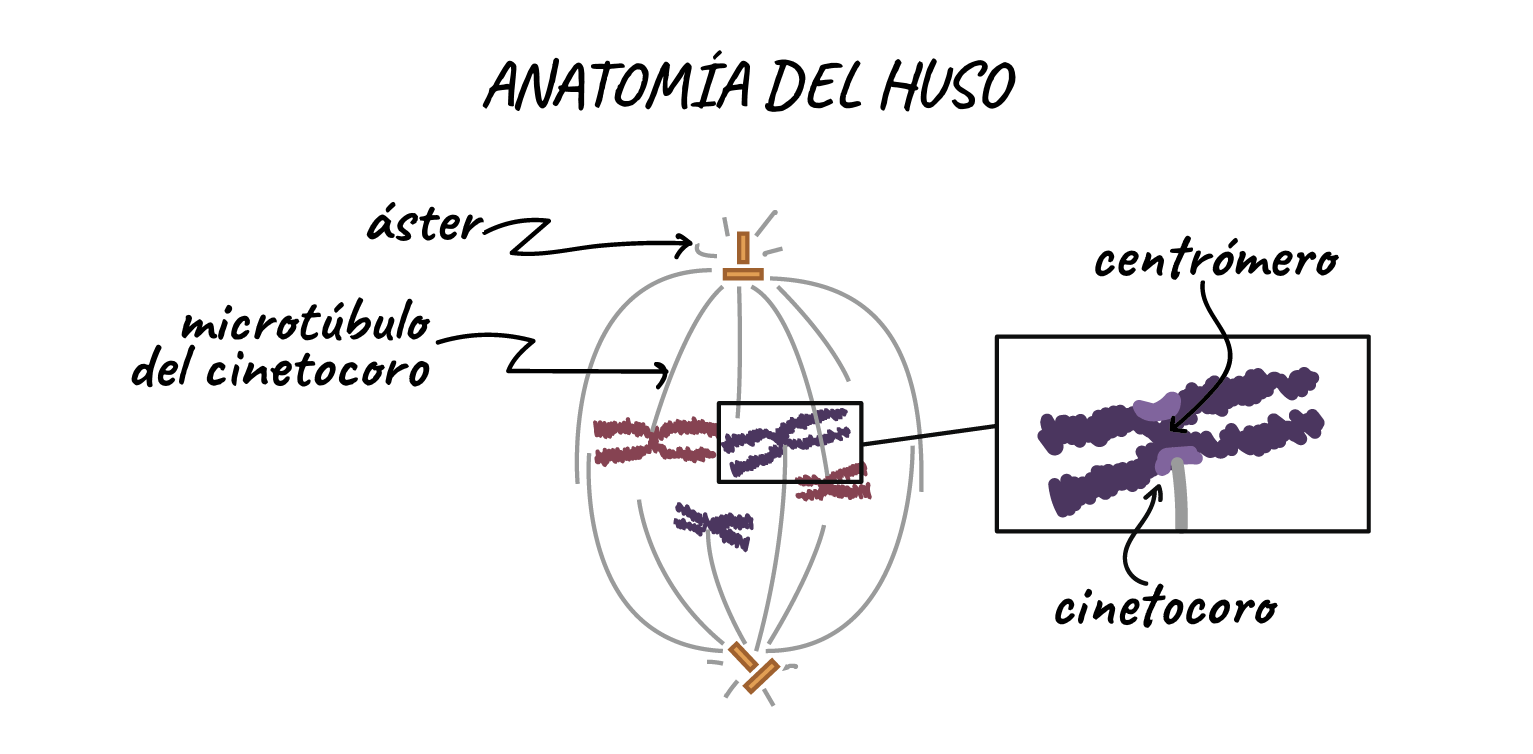
- HUSO MITOTICO
- CILIOS
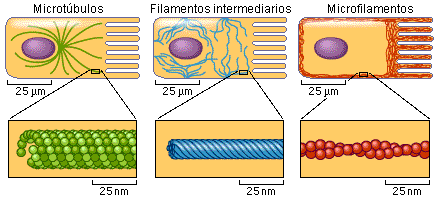
- CITOESQUELETO
- Conjunto de Microfilamentos,
Proporciona el sostén
mecánico.
- Responsable de polaridad celular.,
Otorga a las células musculares
la maquinaria para la contracción.
- Responsable de polaridad celular.,
Otorga a las células musculares
la maquinaria para la contracción.
- proteínas accesorias son
muy importantes para unir
los Microfilamentos.
- Permite a las neuronas
extender los axones y dendritas.
- Permite a las neuronas
extender los axones y dendritas.
- Auto ensamblaje y estructura
- Filamentos de actina
- son dos polímeros helicoidales de la
proteína actina, aparecen como
estructuras flexibles, organizado en una
gran variedad de hases lineales, en redes
bidimensionales y geles tridimensionales
- NUCLEACIÓN
- El GTP favorece el
crecimiento — GDP
acotamiento.
- Se refiere a la formación de 3
monómeros de actina
proporcionandole mayor
estabilidad y actúa como núcleo,
siendo esta una acción reversible
- El GTP favorece el
crecimiento — GDP
acotamiento.
- NUCLEACIÓN
- son dos polímeros helicoidales de la
proteína actina, aparecen como
estructuras flexibles, organizado en una
gran variedad de hases lineales, en redes
bidimensionales y geles tridimensionales
- Microtubulos
- son cilindros largos y huecos formados por la
proteína tubulina,, son rígidos que los
filamentos de actina, normalmente tienen un
extremo unido a un centro organizado de
Microtubulos llamado centrosoma.
- NUCLEACIÓN
- formados por dimero de tubulinas alfa y beta la unión de
estos forman oligómeros que constituyen un núcleo se
alinean ordenadamente mediante enlaces no covalentes,
en filas longitudinales que se denominan protofilamentos
- extremos - formados por alfa-tubulinas
- extremo + formado por beta tubulina*
- son dinámicos
- formados por dimero de tubulinas alfa y beta la unión de
estos forman oligómeros que constituyen un núcleo se
alinean ordenadamente mediante enlaces no covalentes,
en filas longitudinales que se denominan protofilamentos
- NUCLEACIÓN
- son cilindros largos y huecos formados por la
proteína tubulina,, son rígidos que los
filamentos de actina, normalmente tienen un
extremo unido a un centro organizado de
Microtubulos llamado centrosoma.
- Filamentos intermedios
- estructuras parecidas a cuerdas, se localiza debajo la
membrana nuclear interna , otros filamentos
intermedios se extienden a lo largo del citoplasma
proporcionando a la célula resistencia mecánica
- estructuras parecidas a cuerdas, se localiza debajo la
membrana nuclear interna , otros filamentos
intermedios se extienden a lo largo del citoplasma
proporcionando a la célula resistencia mecánica
- Filamentos de actina
- proteínas
accesorias
- Proteínas
motoras
- sirve de molvilidad, contracción y cambios de
formas celulares. Existen 3:
- •Miosina
- •Quinesina
- •Dineina:
- moléculas motoras se diferencian en el tipo de
filamento al que se unen, en el sentido que se
desplazan y en la carga que transportan. Se asocian a
filamentos mediante regiones apicales o ¨cabeza¨
(dominios motores) que unen ATP y lo hidrolizan
- Cabeza: dominio o motor, determina
la identificación de la vía y la
dirección del desplazamiento Colas:
determina el tipo de carga, y la
función biológica de cada proteína.
- Ciclo mecánico químico:
- o Unión al filamento
o Cambio conformacional
o Liberación al filamento
o Generación de fuerza de
unión
- o Unión al filamento
o Cambio conformacional
o Liberación al filamento
o Generación de fuerza de
unión
- Cabeza: dominio o motor, determina
la identificación de la vía y la
dirección del desplazamiento Colas:
determina el tipo de carga, y la
función biológica de cada proteína.
- •Miosina
- sirve de molvilidad, contracción y cambios de
formas celulares. Existen 3:
- Proteinas de
unión o
ligadoras
- conecta los filamentos entre sí y con distinta
características celular de membrana.
- conecta los filamentos entre sí y con distinta
características celular de membrana.
- Proteinas
Reguladoras
- regulan los procesos de alargamiento
(polimerización) y acorta (despolimerización).
- regulan los procesos de alargamiento
(polimerización) y acorta (despolimerización).
- Proteínas
motoras
- Conjunto de Microfilamentos,
Proporciona el sostén
mecánico.
- Cómo regulan las células los
filamentos de su citoesqueleto
- Algunas propiedades de los filamentos
están regulados por modificaciones
covalentes directas de las subunidades
de filamento, pero la mayoría ejerce un
gran conjunto de proteínas accesorias
que se unen a los filamentos
- • En las células animales los
microtubulos se forman a
partir del centrosoma
- • A menudo los filamentos de actina
se nuclean en la membrana
plasmática
- regulada por señales externas permitiendo cambiar
rápidamente su forma y rigidez respondiendo a los
cambios de su entorno, puede estar catalizada por
dos tipos distintos de factores reguladores
- El complejo
ARP
- nuclea el crecimiento de actina a partir
del extremo menos y posibilita de este
modo un alargamiento rápido de los
filamentos por el extremo más.
- nuclea el crecimiento de actina a partir
del extremo menos y posibilita de este
modo un alargamiento rápido de los
filamentos por el extremo más.
- Las
forminas
- de proteínas diméricas, cada subunidad presenta un
lugar de unión para un monómero de actina, el dímero de
formina nuclea la polimerización del filamento, mientras
el nuevo filamento crece, el dímero de formina se
mantiene asociado al extremo más.
- de proteínas diméricas, cada subunidad presenta un
lugar de unión para un monómero de actina, el dímero de
formina nuclea la polimerización del filamento, mientras
el nuevo filamento crece, el dímero de formina se
mantiene asociado al extremo más.
- • El alargamiento de los filamentos está modificado por
proteínas que se unen a las subunidades libres
- Cuando los filamentos han sido nucleados, por lo general se alargan por
adición de subunidades solubles. Conjunto tan abundante contiene
proteínas que se unen a los monómeros de actina e impiden su
polimerización o la hacen menos favorable.
- la más abundante es una pequeña proteína llamada timosina. Los
monómeros de actina unidos a la timiosina se encuentran en un estado
cerrado, en el cual no puede asociarse ni al extremo más ni menos de
los filamentos de actina y tampoco pueden hidrolizar o cambiar el
nucleótido que llevan unido.
- reclutamiento depende de otra proteína de unión al
monómero, la profilina, ésta se une a un monómero de actina
en la cara opuesta de forma que bloquea el lugar al que se
asocia el monómero con el extremo menos. El crecimiento de
los filamentos de actina depende mucho más de la activación
de la profilina para aquellos filamentos cuyos extremos más
están asociados a ciertas forminas.
- • La dinámica de los filamentos puede verse drásticamente alterada por
la unión de proteínas que interaccionan con sus extremos
- • Diferentes tipos de proteínas alteran las propiedades de los extremos de los
microtúbulos que crecen rápidamente
- proteínas que actúan en los extremos de los microtubulos
desempeñan papeles mucho más importantes que los de ser simples
proteínas recubridoras de filamentos. Pueden afectar de forma
dramática la inestabilidad dinámica de los microtubulos, pueden
influenciarla velocidad a la cual los microtubulos cambian de un
estado de crecimiento a un estado de despolimerización, o de un
estado de despolimerización a uno de crecimiento
- • Los filamentos intermedios están empaquetados y entrecruzados
formando hileras compactas.
- grupos de neurofilamentos forman hileras paralelas muy
resistentes que se mantienen unidas por contactos laterales
múltiples, proporcionando estabilidad y fuerza a las largas
expansiones neuronales.
- empaquetar los filamentos intermedios, también los une a los
micrtúbulos, a los haces de filamento de actina y a los filamentos de la
proteína motora miosina II; además permite la unión de los haces de
filamentos intermedios a las estructuras adhesivas de la membrana
plasmática
- •La filamina y la espectrina forman redes de filamentos de actina
- Los geles de actina formados por filamina son necesarios para que
extiendan proyecciones de membrana parecidas a láminas llamadas
lamelipodios que les permite arrastrarse por superficies sólidas.
- descritas disponen de conexiones rectas y rígidas
entre sus dos dominios de unión a los filamento de
actina y tienden a alinear a los filamentos en haces
paralelos.
- • Los elementos del citoesqueleto puede
unirse a la membrana plasmática
- proteínas ERM pueden existir en dos conformaciones, una conformación extendida
activa que oligomeriza y se une a la actina y a una proteína trasmembrana, y una
conformación plegada inactiva, en la cual los extremos N y C-terminales están
empaquetados juntos mediante interacciones intramoleculares. El paso de una
conformación a otra se puede producir por fotosíntesis o por la unión a PIP2
- proteínas ERM pueden existir en dos conformaciones, una conformación extendida
activa que oligomeriza y se une a la actina y a una proteína trasmembrana, y una
conformación plegada inactiva, en la cual los extremos N y C-terminales están
empaquetados juntos mediante interacciones intramoleculares. El paso de una
conformación a otra se puede producir por fotosíntesis o por la unión a PIP2
- Los geles de actina formados por filamina son necesarios para que
extiendan proyecciones de membrana parecidas a láminas llamadas
lamelipodios que les permite arrastrarse por superficies sólidas.
- grupos de neurofilamentos forman hileras paralelas muy
resistentes que se mantienen unidas por contactos laterales
múltiples, proporcionando estabilidad y fuerza a las largas
expansiones neuronales.
- proteínas que actúan en los extremos de los microtubulos
desempeñan papeles mucho más importantes que los de ser simples
proteínas recubridoras de filamentos. Pueden afectar de forma
dramática la inestabilidad dinámica de los microtubulos, pueden
influenciarla velocidad a la cual los microtubulos cambian de un
estado de crecimiento a un estado de despolimerización, o de un
estado de despolimerización a uno de crecimiento
- • Diferentes tipos de proteínas alteran las propiedades de los extremos de los
microtúbulos que crecen rápidamente
- Cuando los filamentos han sido nucleados, por lo general se alargan por
adición de subunidades solubles. Conjunto tan abundante contiene
proteínas que se unen a los monómeros de actina e impiden su
polimerización o la hacen menos favorable.
- El complejo
ARP
- regulada por señales externas permitiendo cambiar
rápidamente su forma y rigidez respondiendo a los
cambios de su entorno, puede estar catalizada por
dos tipos distintos de factores reguladores
- • A menudo los filamentos de actina
se nuclean en la membrana
plasmática
- • En las células animales los
microtubulos se forman a
partir del centrosoma
- Algunas propiedades de los filamentos
están regulados por modificaciones
covalentes directas de las subunidades
de filamento, pero la mayoría ejerce un
gran conjunto de proteínas accesorias
que se unen a los filamentos
Media attachments
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
0 comments
Want to create your own Mind Maps for free with GoConqr? Learn more.