15985378
Description
Mind Map by nicolas penagos, updated more than 1 year ago
|
|
Created by nicolas penagos
about 6 years ago
|
|
DESARROLLO EMBRIONARIO
EQUINODERMOS
- Estos cigotos tienen escaso vitelo
equitativamente distribuido y son
por lo tanto isoleciticos
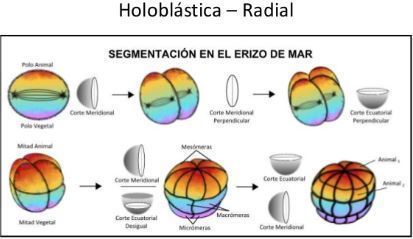
- Segmentación es holoblástica radial
- La primera y la segunda segmentación son meridionales y se disponen perpendiculares entre sí. Es decir, los
surcos de segmentación pasan a través de los polos animal y vegetal
- La tercera segmentación es ecuatorial
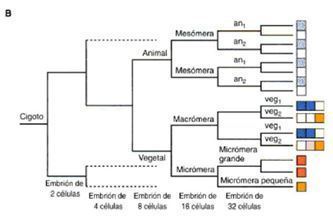
- La cuarta segmentación las cuatro células del piso o nivel animal se dividen meridionalmente en ocho
blastómeras, cada una con el mismo volumen. Estas ocho células son denominadas mesómeras.
- la sexta segmentación. Las células del hemisferio animal se dividen meridionalmente,mientras que
las células vegetales se dividen ecuatorialmente; este patrón es revertido en la séptima división. En
este momento, el embrión es una blástula de 128 células y el patrón de divisiones llega a ser menos
regular.
- Lámina epitelial del blastocele
- Las uniones estrechas unen las blastómeras que antes estaban conectadas laxamente formando una
lámina epitelial continua que rodea completamente el blastocele
- Las uniones estrechas unen las blastómeras que antes estaban conectadas laxamente formando una
lámina epitelial continua que rodea completamente el blastocele
- 4. ¿Cómo el embrión forma la placa vegetal y llega a ser una blástula eclosionada?
- La blástula ciliada comienza a rotar
dentro de la membrana de
fecundación.
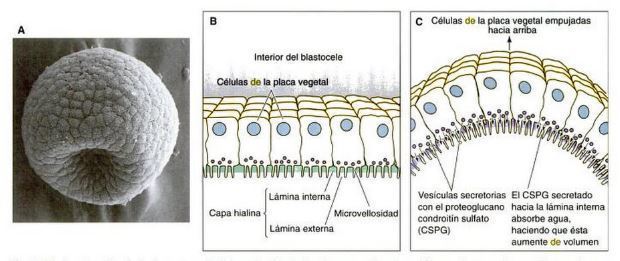
- Las células en el polo vegetal de la blástula
comienzan a engrosarse, formando una
placa vegetal
- Las células en el polo vegetal de la blástula
comienzan a engrosarse, formando una
placa vegetal
- Las células del hemisferio animal sintetizan y secretan
una enzima de eclosión
- La blástula ciliada comienza a rotar
dentro de la membrana de
fecundación.
- La primera y la segunda segmentación son meridionales y se disponen perpendiculares entre sí. Es decir, los
surcos de segmentación pasan a través de los polos animal y vegetal
- Segmentación es holoblástica radial
- DESTINOS CELULARES
- destino es determinado de forma autónoma
- Son las micrómeras esqueletogénicas
- Si son trasplantadas a la región animal de la blástula, no solo sus
descendientes formarán espículas esqueléticas, sino que las
micrómeras trasplantadas alterarán los destino de las células
vecinas al inducir un sitio secundario para la gastrulación.
- Si son trasplantadas a la región animal de la blástula, no solo sus
descendientes formarán espículas esqueléticas, sino que las
micrómeras trasplantadas alterarán los destino de las células
vecinas al inducir un sitio secundario para la gastrulación.
- Son las micrómeras esqueletogénicas
- destino es determinado de forma autónoma
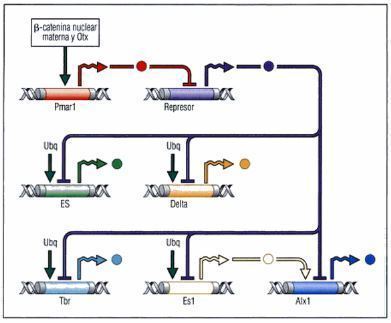
- Vias de señalización
- especificación de las micrómeras
- En primer lugar, durante el desarrollo normal del erizo de mar. la β-catenina se
acumula en los núcleos de las células destinadas a convertirse en endodermo y
mesodermo.
- esta acumulación nuclear parece ser
responsable de la especificación de la mitad
vegetal del embrión.
- esta acumulación nuclear parece ser
responsable de la especificación de la mitad
vegetal del embrión.
- En primer lugar, durante el desarrollo normal del erizo de mar. la β-catenina se
acumula en los núcleos de las células destinadas a convertirse en endodermo y
mesodermo.
- Celulas vegetales
- son especificadas por tres "ondas" de señales
- La primera modera los niveles de β-catenina especificando a las células a ser “endomesodermo”.
- A continuación dos grupo de señales inductivas se emiten
desde las micrómeras. La primera es una señal veg,
temprana" secretada por las micrómeras inmediatamente
despues de su formación en la cuarta segmentación
- La vía Notch hace que estas células lleguen a ser
mesénquima secundario en lugar de endodermo.
- Finalmente, Wnt8 parece ser producido por la células del
endodermo (es decir, aquellas célula del endomesodermo que
no reciben la señal Delta).
- La primera modera los niveles de β-catenina especificando a las células a ser “endomesodermo”.
- son especificadas por tres "ondas" de señales
- especificación de las micrómeras
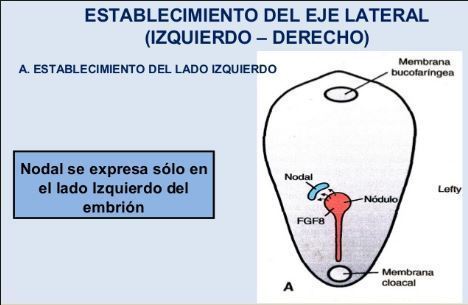
- Ejes corporales
- En la mayoría de los erizos de mar, los eje dorsoventral e
izquierda-derecha son especificados después de la
fecundación, pero el modo de su especificación no es bien
entendido.
- El eje oral-aboral (ventral-dorsal) de los embriones
de erizo de mar en general es definido por el
primer plano de segmentación.
- Curiosamente, en los erizos de mar que evitan entrar en el estadio
larval para desarrollarse directamente a juveniles, el eje
dorsoventral es especificado maternamente en el citoplasma del
óvulo.
- Curiosamente, en los erizos de mar que evitan entrar en el estadio
larval para desarrollarse directamente a juveniles, el eje
dorsoventral es especificado maternamente en el citoplasma del
óvulo.
- El eje oral-aboral (ventral-dorsal) de los embriones
de erizo de mar en general es definido por el
primer plano de segmentación.
- En la mayoría de los erizos de mar, los eje dorsoventral e
izquierda-derecha son especificados después de la
fecundación, pero el modo de su especificación no es bien
entendido.
- Gastrulación
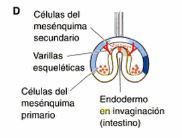
- lngresión: Migración de células individuales de la capa superficial hacia el
interior del embrión. Las células se convierten en mesenquimáticas
(separadas) y migran en forma independiente.
- Invaginación del arquenteron
- Se detiene la invaginación, la region invaginada se llama
arquenteron, y se origina el blsatoporo en el polo vegetal
- La celulas del polo vegetal adoptan la forma de botella
- La capa hialina se incurva hacia adentro
- La capa hialina se incurva hacia adentro
- La celulas del polo vegetal adoptan la forma de botella
- Extensión convergente
- Pausa breve despues de la invaginación inicial
- Extensión exagerada
- Rudimiento ancho y grueso del intestino
se transforma en un tubo delgado y fino
- Para llevarlo a cabo se reorganiza el arquenteron
- Para llevarlo a cabo se reorganiza el arquenteron
- Rudimiento ancho y grueso del intestino
se transforma en un tubo delgado y fino
- Extensión exagerada
- Pausa breve despues de la invaginación inicial
- Se detiene la invaginación, la region invaginada se llama
arquenteron, y se origina el blsatoporo en el polo vegetal
- lngresión: Migración de células individuales de la capa superficial hacia el
interior del embrión. Las células se convierten en mesenquimáticas
(separadas) y migran en forma independiente.
- Matriz extracelular
- primer movimiento de gastrulación
- aumento de afinidad por la lamina basal
- las micromeras liberan su adhesiones a la capa hialina
- Retraccion por la lamina basal
- Migración al blastocele
- Migración al blastocele
- Retraccion por la lamina basal
- las micromeras liberan su adhesiones a la capa hialina
- aumento de afinidad por la lamina basal
- primer movimiento de gastrulación
Media attachments
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Want to create your own Mind Maps for free with GoConqr? Learn more.