21037751
Description
Mind Map by Jacces Luna, updated more than 1 year ago
|
|
Created by Jacces Luna
almost 5 years ago
|
|
Curva de disociación
de oxihemoglobina
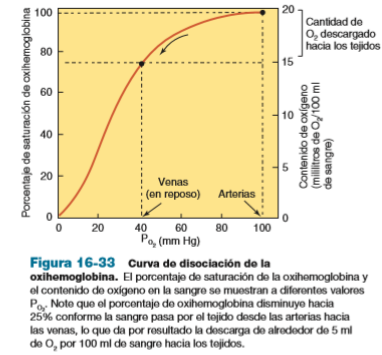
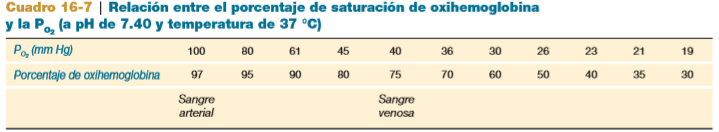
- La sangre en las arterias sistémicas,
a una PO2 de 100 mm Hg, tiene un
porcentaje de saturación de
oxihemoglobina de 97% (lo que
significa que 97% de la
hemoglobina está en la forma de
oxihemoglobina).
- Esta sangre es
suministrada a los
capilares sistémicos, donde
el oxígeno se difunde hacia
las células y se consume en
la respiración aeróbica.
- De este modo, la sangre que sale en las venas
sistémicas está reducida en oxígeno; tiene una PO2
de alrededor de 40 mm Hg, y un porcentaje de
saturación de oxihemoglobina de aproximadamente
75% cuando una persona está en reposo.
- En otras palabras, la sangre
que entra a los tejidos
contiene 20 ml de O2 por 100
ml de sangre, y la que sale de
los tejidos contiene 15.5 ml de
O2 por 100 ml de sangre
- Así, 22%, o 4.5 ml de
O2 de los 20 ml de
O2 por 100 ml de
sangre, se descarga
hacia los tejidos.
- Una ilustración gráfica del
porcentaje de saturación de
oxihemoglobina a diferentes
valores de PO2 se llama una
curva de disociación de
oxihemoglobina
- Los valores en este gráfico se
obtienen al sujetar muestras de
sangre in vitro a diferentes
presiones parciales de oxígeno
- la PO2 arterial y venosa, y
el porcentaje de
saturación de
oxihemoglobina en
reposo.
- La cantidad de oxihemoglobina
relativamente grande que queda en la
sangre venosa en reposo sirve como
una reserva de oxígeno.
- Si una persona deja de respirar, una
reserva suficiente de oxígeno en la
sangre mantendrá el encéfalo y el
corazón vivos durante alrededor de 4 a 5
min sin el uso de técnicas de
reanimación cardiopulmonar
(cardiopulmonary resuscitation [CPR]).
- Este aporte de reserva de
oxígeno también puede utilizarse
cuando los requerimientos de
oxígeno de un tejido aumentan,
como en los músculos que están
haciendo ejercicio.
- La curva de disociación de la oxihemoglobina
tiene forma de S, o sigmoidea. El hecho de que
es relativamente plana a valores de PO2 altos
indica que los cambios de la PO2 dentro de este
rango tienen poco efecto sobre la reacción de
carga. Por ejemplo, sería necesario ascender
hasta a 3 050 m (10 000 pies) antes de que la
saturación de oxihemoglobina de la sangre
arterial disminuyera desde 97 hasta 93%.
- A elevaciones más
comunes, el porcentaje de
saturación de
oxihemoglobina no sería
significativamente distinto
del valor de 97% al nivel del
mar.
- Empero, en la parte empinada de la
curva sigmoidea, cambios pequeños de
los valores de PO2 producen grandes
diferencias del porcentaje de
saturación. Una disminución de la PO2
venosa desde 40 mm Hg hasta 30 mm
Hg, como podría ocurrir durante el
ejercicio leve, corresponde a un cambio
del porcentaje de saturación desde 75
hasta 58%.
- Dado que el porcentaje de saturación arterial por lo
general aún es de 97% durante el ejercicio, el
porcentaje de saturación venosa disminuido indica
que se ha descargado más oxígeno hacia los tejidos.
La diferencia entre los porcentajes de saturación
arterial y venoso indica el porcentaje de descarga. En
el ejemplo precedente, 97% − 75% = 22% de descarga
en reposo, y 97% − 58% = 39% de descarga durante el
ejercicio leve. Durante el ejercicio más pesado, la
PO2 venosa puede disminuir a 20 mm Hg o menos, lo
que indica un porcentaje de descarga de alrededor
de 80%
- Dado que el porcentaje de saturación arterial por lo
general aún es de 97% durante el ejercicio, el
porcentaje de saturación venosa disminuido indica
que se ha descargado más oxígeno hacia los tejidos.
La diferencia entre los porcentajes de saturación
arterial y venoso indica el porcentaje de descarga. En
el ejemplo precedente, 97% − 75% = 22% de descarga
en reposo, y 97% − 58% = 39% de descarga durante el
ejercicio leve. Durante el ejercicio más pesado, la
PO2 venosa puede disminuir a 20 mm Hg o menos, lo
que indica un porcentaje de descarga de alrededor
de 80%
- Empero, en la parte empinada de la
curva sigmoidea, cambios pequeños de
los valores de PO2 producen grandes
diferencias del porcentaje de
saturación. Una disminución de la PO2
venosa desde 40 mm Hg hasta 30 mm
Hg, como podría ocurrir durante el
ejercicio leve, corresponde a un cambio
del porcentaje de saturación desde 75
hasta 58%.
- A elevaciones más
comunes, el porcentaje de
saturación de
oxihemoglobina no sería
significativamente distinto
del valor de 97% al nivel del
mar.
- La curva de disociación de la oxihemoglobina
tiene forma de S, o sigmoidea. El hecho de que
es relativamente plana a valores de PO2 altos
indica que los cambios de la PO2 dentro de este
rango tienen poco efecto sobre la reacción de
carga. Por ejemplo, sería necesario ascender
hasta a 3 050 m (10 000 pies) antes de que la
saturación de oxihemoglobina de la sangre
arterial disminuyera desde 97 hasta 93%.
- Este aporte de reserva de
oxígeno también puede utilizarse
cuando los requerimientos de
oxígeno de un tejido aumentan,
como en los músculos que están
haciendo ejercicio.
- Si una persona deja de respirar, una
reserva suficiente de oxígeno en la
sangre mantendrá el encéfalo y el
corazón vivos durante alrededor de 4 a 5
min sin el uso de técnicas de
reanimación cardiopulmonar
(cardiopulmonary resuscitation [CPR]).
- La cantidad de oxihemoglobina
relativamente grande que queda en la
sangre venosa en reposo sirve como
una reserva de oxígeno.
- la PO2 arterial y venosa, y
el porcentaje de
saturación de
oxihemoglobina en
reposo.
- Los valores en este gráfico se
obtienen al sujetar muestras de
sangre in vitro a diferentes
presiones parciales de oxígeno
- Una ilustración gráfica del
porcentaje de saturación de
oxihemoglobina a diferentes
valores de PO2 se llama una
curva de disociación de
oxihemoglobina
- Así, 22%, o 4.5 ml de
O2 de los 20 ml de
O2 por 100 ml de
sangre, se descarga
hacia los tejidos.
- En otras palabras, la sangre
que entra a los tejidos
contiene 20 ml de O2 por 100
ml de sangre, y la que sale de
los tejidos contiene 15.5 ml de
O2 por 100 ml de sangre
- De este modo, la sangre que sale en las venas
sistémicas está reducida en oxígeno; tiene una PO2
de alrededor de 40 mm Hg, y un porcentaje de
saturación de oxihemoglobina de aproximadamente
75% cuando una persona está en reposo.
- Esta sangre es
suministrada a los
capilares sistémicos, donde
el oxígeno se difunde hacia
las células y se consume en
la respiración aeróbica.
- Efecto del pH y la
temperatura
sobre el
transporte de
oxígeno
- Además de los cambios de la PO2, las reacciones
de carga y descarga están influidas por cambios
de la afinidad (la fuerza del enlace) de la
hemoglobina por el oxígeno. Tales cambios
aseguran que los músculos esqueléticos activos
recibirán más oxígeno desde la sangre que el que
reciben en reposo. Esto ocurre como resultado del
pH disminuido y la temperatura aumentada en
los músculos que están haciendo ejercicio.
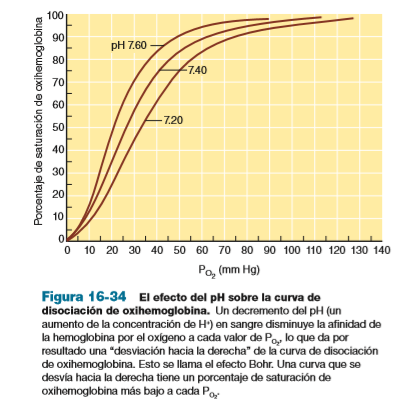
- La afinidad disminuye
cuando el pH está
disminuido, y aumenta
cuando el pH está
aumentado; esto se llama el
efecto Bohr.
- Cuando la afinidad de la
hemoglobina por el oxígeno se
reduce, hay un poco menos de
carga de la sangre con oxígeno
en los pulmones, pero mayor
descarga de oxígeno en los
tejidos.
- El efecto neto es que los tejidos
reciben más oxígeno cuando el pH
de la sangre está disminuido
- Dado que el dióxido de
carbono puede disminuir el
pH (mediante la formación
de ácido carbónico), el efecto
Bohr ayuda a proporcionar
más oxígeno a los tejidos
cuando su producción de
dióxido de carbono está
aumentada por un
metabolismo más rápido.
- Cuando se analizan las curvas de
disociación de la oxihemoglobina
graficadas a diferentes valores
de pH, puede observarse que la
curva de disociación se desvía
hacia la derecha por una
disminución del pH, y hacia la
izquierda por un aumento del pH
- Si se calcula el porcentaje de descarga (al
restar el porcentaje de saturación de
oxihemoglobina para sangre arterial y
venosa), se observará que una desviación
de la curva hacia la derecha indica una
mayor descarga de oxígeno. Por el
contrario, una desviación hacia la
izquierda indica menos descarga pero
carga de un poco más de oxígeno en los
pulmones.
- Si se calcula el porcentaje de descarga (al
restar el porcentaje de saturación de
oxihemoglobina para sangre arterial y
venosa), se observará que una desviación
de la curva hacia la derecha indica una
mayor descarga de oxígeno. Por el
contrario, una desviación hacia la
izquierda indica menos descarga pero
carga de un poco más de oxígeno en los
pulmones.
- Cuando se analizan las curvas de
disociación de la oxihemoglobina
graficadas a diferentes valores
de pH, puede observarse que la
curva de disociación se desvía
hacia la derecha por una
disminución del pH, y hacia la
izquierda por un aumento del pH
- Dado que el dióxido de
carbono puede disminuir el
pH (mediante la formación
de ácido carbónico), el efecto
Bohr ayuda a proporcionar
más oxígeno a los tejidos
cuando su producción de
dióxido de carbono está
aumentada por un
metabolismo más rápido.
- El efecto neto es que los tejidos
reciben más oxígeno cuando el pH
de la sangre está disminuido
- Cuando la afinidad de la
hemoglobina por el oxígeno se
reduce, hay un poco menos de
carga de la sangre con oxígeno
en los pulmones, pero mayor
descarga de oxígeno en los
tejidos.
- La afinidad disminuye
cuando el pH está
disminuido, y aumenta
cuando el pH está
aumentado; esto se llama el
efecto Bohr.
- Además de los cambios de la PO2, las reacciones
de carga y descarga están influidas por cambios
de la afinidad (la fuerza del enlace) de la
hemoglobina por el oxígeno. Tales cambios
aseguran que los músculos esqueléticos activos
recibirán más oxígeno desde la sangre que el que
reciben en reposo. Esto ocurre como resultado del
pH disminuido y la temperatura aumentada en
los músculos que están haciendo ejercicio.
- Efecto del 2,3-DPG
sobre el
transporte de
oxígeno
- Los eritrocitos maduros carecen tanto de
núcleo como de mitocondrias. Sin
mitocondrias no pueden respirar de manera
aeróbica; ¡las células mismas que transportan
oxígeno son las únicas células en el cuerpo
que no pueden usarlo!
- De este modo, los eritrocitos deben
obtener energía por medio del
metabolismo anaeróbico de glucosa. En un
cierto punto de la vía glucolítica, ocurre
una “reacción colateral” en los eritrocitos
que da por resultado un producto singular:
el ácido 2,3-difosfoglicérico (2,3-DPG).
- La enzima que produce el 2,3-DPG es inhibida por la
oxihemoglobina. En consecuencia, cuando la
concentración de oxihemoglobina está disminuida, la
producción de 2,3-DPG está aumentada. Este
incremento de la producción de 2,3-DPG puede ocurrir
cuando la concentración total de hemoglobina es baja
(en presencia de anemia) o cuando la PO2 es baja (a una
altitud elevada
- El enlace de 2,3-DPG con desoxihemoglobina hace que esta
última sea más estable. Por tanto, una proporción más alta
de la oxihemoglobina se convertirá en desoxihemoglobina
por la descarga de su oxígeno. Así, una concentración
aumentada de 2,3-DPG en los eritrocitos aumenta la
descarga de oxígeno y desvía la curva de disociación de
oxihemoglobina hacia la derecha.
- El enlace de 2,3-DPG con desoxihemoglobina hace que esta
última sea más estable. Por tanto, una proporción más alta
de la oxihemoglobina se convertirá en desoxihemoglobina
por la descarga de su oxígeno. Así, una concentración
aumentada de 2,3-DPG en los eritrocitos aumenta la
descarga de oxígeno y desvía la curva de disociación de
oxihemoglobina hacia la derecha.
- La enzima que produce el 2,3-DPG es inhibida por la
oxihemoglobina. En consecuencia, cuando la
concentración de oxihemoglobina está disminuida, la
producción de 2,3-DPG está aumentada. Este
incremento de la producción de 2,3-DPG puede ocurrir
cuando la concentración total de hemoglobina es baja
(en presencia de anemia) o cuando la PO2 es baja (a una
altitud elevada
- De este modo, los eritrocitos deben
obtener energía por medio del
metabolismo anaeróbico de glucosa. En un
cierto punto de la vía glucolítica, ocurre
una “reacción colateral” en los eritrocitos
que da por resultado un producto singular:
el ácido 2,3-difosfoglicérico (2,3-DPG).
- Los eritrocitos maduros carecen tanto de
núcleo como de mitocondrias. Sin
mitocondrias no pueden respirar de manera
aeróbica; ¡las células mismas que transportan
oxígeno son las únicas células en el cuerpo
que no pueden usarlo!
Media attachments
{kind=link}
{kind=link}
{kind=link}
Want to create your own Mind Maps for free with GoConqr? Learn more.