10659520
Description
Mind Map by Tara van Zyl, updated more than 1 year ago
|
|
Created by Tara van Zyl
over 7 years ago
|
|
Visual Brain
Part I
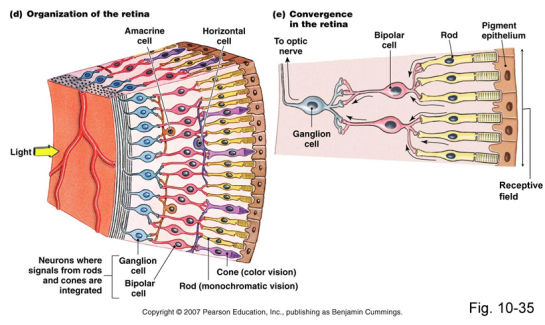
- Retinal Structure
- Inner layer of eye- when light hits,
initiates chemical and electrical events
that trigger nerve impulses. Contains
10 layers, 3 of the most important
being: (*also, astrocytes- Muller cells;
interneurons- horizontal cells)
- PHOTORECEPTOR CELL LAYER
electromagnetic energy =
neural signal
- 100,000,000
- RODS= SCOTOPIC VISION
(LOW-LIGHT LEVELS);
CONES= COLOUR VISION
& ACUITY
- FOVEAL VISION- CENTRAL 1 DEGREE OF VISUAL
ANGLE; MACULAR VISION- CENTRAL 5
DEGREES.. Beyond this is peripheral.
- FOVEAL VISION- CENTRAL 1 DEGREE OF VISUAL
ANGLE; MACULAR VISION- CENTRAL 5
DEGREES.. Beyond this is peripheral.
- Rods and cones produce graded
responses- light=hyperpolarisation and
release of neurotransmitters
- Maximal stimulation of the receptors is
determined by the spectral sensitivity of
the photopigment, located on the outer
segment of the receptor (Marks, 1965).
There are more of these in cones
suggesting they are more involved in
processing wavelength/colour.
- Maximal stimulation of the receptors is
determined by the spectral sensitivity of
the photopigment, located on the outer
segment of the receptor (Marks, 1965).
There are more of these in cones
suggesting they are more involved in
processing wavelength/colour.
- 100,000,000
- BIPOLAR CELL LAYER
- 10,000,000
- 10,000,000
- GANGLION CELL LAYER
- 1,250,000
- Ganglion cells carry info
along the optic nerve,
which is made up of axons
from all retinal ganglion
cells & glial cells.
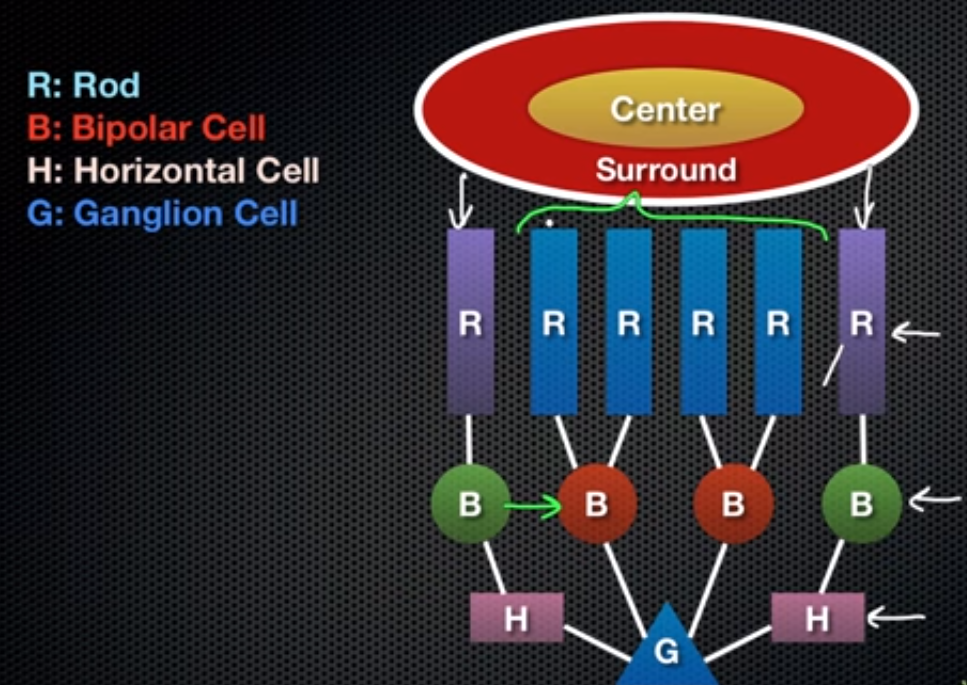
- RECEPTIVE FIELDS OF GANGLION CELLS: area of the retina from which it receives input. Their receptive
fields have a centre-surround configuration- 2 TYPES: ON centre/OFF surround // OFF centre/ON
surround i.e. photoreceptors-bipolar-ganglion is centre; the addition of a horizontal cell in this link
means it is no longer direction and now considered the surround. Depolarisation of centre if on and
hyperpolarisation of surround if off (ON centre: impulses sent during depolarisation; OFF surround:
post-inhibitory rebound impulses sent after hyperpolarisation).
- RECEPTIVE FIELD SIZE: Increasing eccentricity
(deviation from fovea) = increasing receptive field
size. Closer to fovea means greater acuity (smaller
receptive fields) 0.01mm to 0.5mm. Cells respond
best to matching stimuli i.e. small stimuli - small
receptive fields, which suggests object size analysis
may start in the retina.
- RECEPTIVE FIELD SIZE: Increasing eccentricity
(deviation from fovea) = increasing receptive field
size. Closer to fovea means greater acuity (smaller
receptive fields) 0.01mm to 0.5mm. Cells respond
best to matching stimuli i.e. small stimuli - small
receptive fields, which suggests object size analysis
may start in the retina.
- Classification: as well as centre/surround & receptive
field size: A) M cells (magnocellular) B) P cells
(parvocellular) C) K cells (koniocellular)
- P CELLS: small; in fovea; high spatial
resolution; low temporal resolution; low
contrast sensitivity; responds to colour;
conduction velocity slow; 80% of GC.
- M CELLS: large; periphery; all qualities
opposite to P cells, 10% of GC.
- P CELLS: small; in fovea; high spatial
resolution; low temporal resolution; low
contrast sensitivity; responds to colour;
conduction velocity slow; 80% of GC.
- RECEPTIVE FIELDS OF GANGLION CELLS: area of the retina from which it receives input. Their receptive
fields have a centre-surround configuration- 2 TYPES: ON centre/OFF surround // OFF centre/ON
surround i.e. photoreceptors-bipolar-ganglion is centre; the addition of a horizontal cell in this link
means it is no longer direction and now considered the surround. Depolarisation of centre if on and
hyperpolarisation of surround if off (ON centre: impulses sent during depolarisation; OFF surround:
post-inhibitory rebound impulses sent after hyperpolarisation).
- 1,250,000
- PHOTORECEPTOR CELL LAYER
electromagnetic energy =
neural signal
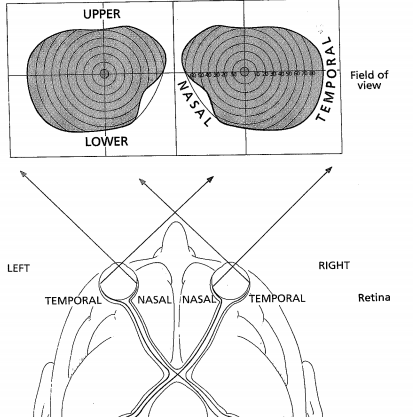
- CURVATURE OF THE EYE: Nasal retina registers
temporal field & vice versa; lower retina registers
upper visual field & vice versa. Thus, left hemi-field is
observed by right temporal & left nasal.
- Inner layer of eye- when light hits,
initiates chemical and electrical events
that trigger nerve impulses. Contains
10 layers, 3 of the most important
being: (*also, astrocytes- Muller cells;
interneurons- horizontal cells)
- Moving from Retina to LGN
- THE OPTIC NERVE: each GC axon acquires
myelin coating- insulates and increases
speed.
- Following the Optic Nerve is the OPTIC CHIASM- here, decrossation
occurs. The RETINA can be subdivided into 4 segments: NASAL,
TEMPORAL, UPPER, LOWER. It can also be subdivided into PERIPHERAL
and CENTRAL.
- Optic nerves for nasal retina of a specific eye cross over to the opposite hemisphere for
processing in the LGN; contrarily, the optic nerves for the temporal retina of that eye do not
cross over. Contralateral/ipsilateral.
- Visual pathway after this becomes known as the
OPTIC TRACT, which informs the LGN in the cerebral
hemisphere- 6 layered structure
- Inputs to the LGN are segregated: fibres coming from ipsilateral eye terminate in layers
5, 3 & 2; contralateral eye terminate 6, 4 & 1. Point to point projection means ^ acuity,
but also each eye is represented twice.
- Points on layers correspond highly to points on retina and this is the same for
each eye i.e. same point different layer, corresponds to eye.
- These 6 layers are made up of cells- the upper 4 being parvocellular cells and the
lower 2 being magnocellular cells. They correspond to the matching GC type and
are sourced from rods (m) and cones (p)
- *CRITICISM/HISTORY: Henschen erroneously
believed p cells code only for colour & that m
cells code only for light and that this was
implicated in their size.
- LGN cell axons travel in optic radiation to terminate at the PVC.
- LGN cell axons travel in optic radiation to terminate at the PVC.
- *CRITICISM/HISTORY: Henschen erroneously
believed p cells code only for colour & that m
cells code only for light and that this was
implicated in their size.
- These 6 layers are made up of cells- the upper 4 being parvocellular cells and the
lower 2 being magnocellular cells. They correspond to the matching GC type and
are sourced from rods (m) and cones (p)
- Points on layers correspond highly to points on retina and this is the same for
each eye i.e. same point different layer, corresponds to eye.
- Inputs to the LGN are segregated: fibres coming from ipsilateral eye terminate in layers
5, 3 & 2; contralateral eye terminate 6, 4 & 1. Point to point projection means ^ acuity,
but also each eye is represented twice.
- As well as going to the
LGN, the retina
connects to the
superior colliculus &
pulvinar. This is key to
understanding certain
types of blindness e.g.
blindsight
- Visual pathway after this becomes known as the
OPTIC TRACT, which informs the LGN in the cerebral
hemisphere- 6 layered structure
- Optic nerves for nasal retina of a specific eye cross over to the opposite hemisphere for
processing in the LGN; contrarily, the optic nerves for the temporal retina of that eye do not
cross over. Contralateral/ipsilateral.
- Following the Optic Nerve is the OPTIC CHIASM- here, decrossation
occurs. The RETINA can be subdivided into 4 segments: NASAL,
TEMPORAL, UPPER, LOWER. It can also be subdivided into PERIPHERAL
and CENTRAL.
- THE OPTIC NERVE: each GC axon acquires
myelin coating- insulates and increases
speed.
- CITATIONS: Parallel processing and GC receptive fields:
Celesia & DeMarco (1994). Retina, LGN, V1 & V2: Zeki
(1993), Prasad & Galetta (2011).
- The Primary & Secondary Visual Cortex
- "...parallel processing is a hallmark of sensory coding..." Erickson (1974)
- Radiation terminates in the occipital lobes- known as area V1
(each hemisphere has visual cortex).
- Cortex receives signals from contralateral hemifield; adjacent
retinal points accounted for on topographical map (RETINOTOPIC).
- Cytoarchitectonic staining (RNA within cells stained and thus highlighted)
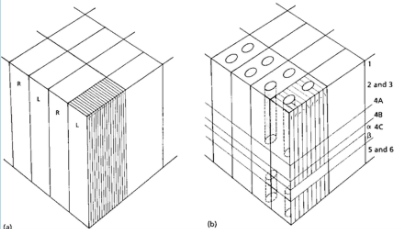
reveals a stratified structure with 6 layers. Layer 4 is larger than usual.
- Layer 4 is comprised of 4 sublayers. The LGN projects
to these layers. Cells in them are monocularly driven
(other layers- binocularly driven)
- MAGNOCELLULAR ROUTE: from LGN to 4Cα
to 4B to other cortical areas.
- PARVOCELLULAR ROUTE: from LGN to 4A
to 4Cβ to 2 & 3 to other cortical areas
- Lower half of visual field is mapped onto upper calcarine cortex, and vice-versa (Zeki, 1993); central
vision at occipital pole (*previous Henschen assumption that it had been anterior)
- CORTICAL MAGNIFICATION: a disproportionate number of cells in the V1 deal with information
from the central retina (80% to 10 degrees) - receptive fields and eccentricity.
- AREA V2- prestriate cortex. Also indirect link between V1 and V3-5. Mapped as V1 is.
- Has a thin stripe (connections from blobs in V1, wavelength) and thick stripe (from 4B,
orientation *direction), and interstripe (interblobs- orientation only)
- THIN STRIPES TO V4
- iNTERSTRIPES TO V4
- THICK STRIPES - ORIENTATION CELLS TO V3;
ORIENTATION & DIRECTION CELLS TO V5
- MOTION PATHWAY- M GC CELLS IN RET -- M LAYERS IN
LGN -- LAYER 4B IN V1 (ORIENTATION & MOTION) --
THICK STRIPES IN V2 -- AREA V5
- DYNAMIC FORM PATHWAY- M GC CELLS IN RET --
M LAYERS IN LGN -- LAYER 4B ORINETATION IN V1
-- THICK STRIPES IN V2 -- AREA V3.
- COLOUR PATHWAY- P GC CELLS IN RET -- P LAYERS
IN LGN -- 2 & 3 IN V1 BLOB CELLS WAVELENGTH --
THIN STRIPES V2 -- AREA V4
- FORM & COLOUR PATHWAY- P GC CELLS IN RET -- P
LAYERS IN LGN -- LAYERS 2 & 3 V1 INTERBLOB
ORIENTATION -- INTERSTRIPES IN V2 -- AREA V4
- FORM & COLOUR PATHWAY- P GC CELLS IN RET -- P
LAYERS IN LGN -- LAYERS 2 & 3 V1 INTERBLOB
ORIENTATION -- INTERSTRIPES IN V2 -- AREA V4
- COLOUR PATHWAY- P GC CELLS IN RET -- P LAYERS
IN LGN -- 2 & 3 IN V1 BLOB CELLS WAVELENGTH --
THIN STRIPES V2 -- AREA V4
- DYNAMIC FORM PATHWAY- M GC CELLS IN RET --
M LAYERS IN LGN -- LAYER 4B ORINETATION IN V1
-- THICK STRIPES IN V2 -- AREA V3.
- THIN STRIPES TO V4
- Has a thin stripe (connections from blobs in V1, wavelength) and thick stripe (from 4B,
orientation *direction), and interstripe (interblobs- orientation only)
- AREA V2- prestriate cortex. Also indirect link between V1 and V3-5. Mapped as V1 is.
- Layer 4 is comprised of 4 sublayers. The LGN projects
to these layers. Cells in them are monocularly driven
(other layers- binocularly driven)
- Cytoarchitectonic staining (RNA within cells stained and thus highlighted)
reveals a stratified structure with 6 layers. Layer 4 is larger than usual.
- PROPERTY 1) ORIENTATION SELECTIVITY (Hubel & Wiesel,1981): most
cells sensitive, unlike RET and LGN. Each has own preferred orientation-
elicit optimal response. Electrode and oscilloscope record heightened
activity when exposed to preferred ort.
- In line with receptive field shape, which is a column as opposed to circular- SIMPLE
relationship between field layout and preferred orientation. Multiple LGN to 1 simple.
- COMPLEX cells - on and off ones are not as clear.
Do NOT respond to static/stationary stimuli-
moving & at optimal ort, direction.
- HYPERCOMPLEX cells: responds to luminance in particular
part of receptive field, of specific length (end-stopping- inhibitory flanks).
- HYPERCOMPLEX cells: responds to luminance in particular
part of receptive field, of specific length (end-stopping- inhibitory flanks).
- COMPLEX cells - on and off ones are not as clear.
Do NOT respond to static/stationary stimuli-
moving & at optimal ort, direction.
- In line with receptive field shape, which is a column as opposed to circular- SIMPLE
relationship between field layout and preferred orientation. Multiple LGN to 1 simple.
- PROPERTY 2) SIZE SELECTIVITY: As with LGN &
RET, cells receiving input from fovea = small RF,
periphery = large RF. Unlike LGN & RET, also
has orientation selectivity.
- The cytochrome oxidase method (Wong-Riley, 1979) reveals an even more
complex anatomical architecture; staining sections for the metabolic
enzyme cytochrome oxidase: POLKA DOT PATTERN layers 2 & 3- DARK
BLOBS (wavelength) and LIGHT INTERBLOBS (orientation).
- N.B. Blobs with
same wavelength
selectivity connect
- OTHER ARCHITECTURAL METHODS: A) Cortical lesions: when destroyed fibres
regenerate and as a result take up more silver (become more argyrophilic) -
areas showing more silver denotes connection to the damaged area.
- B) Radioactive amino acids: injection- AAs taken up by cells in site, then
transported to axon terminals, thus highlighting them.
- C) Enzyme horseradish peroxidase (HRP): injected at
suspected termination site. HRP is transported
retroactively to cell body- thus revealing layer location too.
Can be combined with lectin wheatgerm glutinin (WGA) to
show both prograde and retrograde projections (Zeki, 1993).
- C) Enzyme horseradish peroxidase (HRP): injected at
suspected termination site. HRP is transported
retroactively to cell body- thus revealing layer location too.
Can be combined with lectin wheatgerm glutinin (WGA) to
show both prograde and retrograde projections (Zeki, 1993).
- B) Radioactive amino acids: injection- AAs taken up by cells in site, then
transported to axon terminals, thus highlighting them.
- N.B. Blobs with
same wavelength
selectivity connect
- The cytochrome oxidase method (Wong-Riley, 1979) reveals an even more
complex anatomical architecture; staining sections for the metabolic
enzyme cytochrome oxidase: POLKA DOT PATTERN layers 2 & 3- DARK
BLOBS (wavelength) and LIGHT INTERBLOBS (orientation).
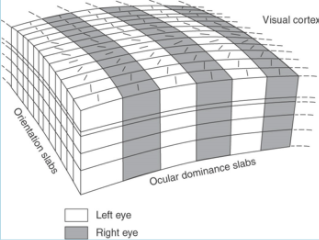
- PROPERTY 3) BINOCULARITY: Layer 4 has
monocular segregation; unlike LGN, rest
of V1 has BS. Individual cells respond to
both eyes but usually to one better than
the other - ocular dominance.
- Preferential response is one of two properties, other- relative
positions of RF specify location in space that will excite cell.
- Functional Architecture-
Each hypercolumn analyses
orientation, motion
direction, binocularity and
size within a local region of
the retina.
- Functional Architecture-
Each hypercolumn analyses
orientation, motion
direction, binocularity and
size within a local region of
the retina.
- Preferential response is one of two properties, other- relative
positions of RF specify location in space that will excite cell.
- PROPERTY 4) MOTION DETECTION:
Shipp & Zeki (1989)- neurons in 4B
sensitive to motion direction- and
they also form connections.
- Cortex receives signals from contralateral hemifield; adjacent
retinal points accounted for on topographical map (RETINOTOPIC).
- Radiation terminates in the occipital lobes- known as area V1
(each hemisphere has visual cortex).
- "...parallel processing is a hallmark of sensory coding..." Erickson (1974)
- DAMAGE
- Retina + optic nerve = total blindness
- Beyond chiasm = contralateral field of view
blind i.e. homonymoushemianopia.
- Occipital pole receives dual blood supply from posterior and middle
cerebral arteries (rest only posterior) = macular sparing
- Upper calcerine sulcus lip -> lower visual
field and vice versa = quadrantanopia
- Small area of damage
in V1 = scotoma,
determined by
perimetry.
- Retina + optic nerve = total blindness
Media attachments
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Want to create your own Mind Maps for free with GoConqr? Learn more.