1152930
Description
Mind Map by Kristi Brogden, updated more than 1 year ago
|
|
Created by Kristi Brogden
over 11 years ago
|
|
DNA replication
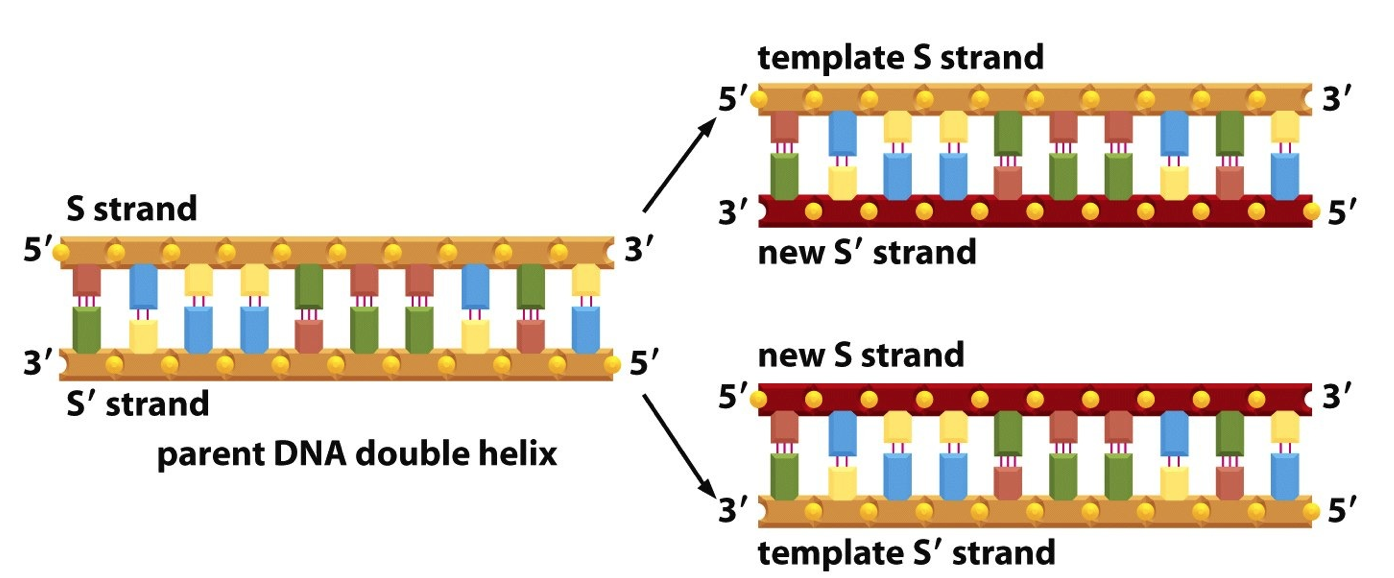
- Semi-conservative
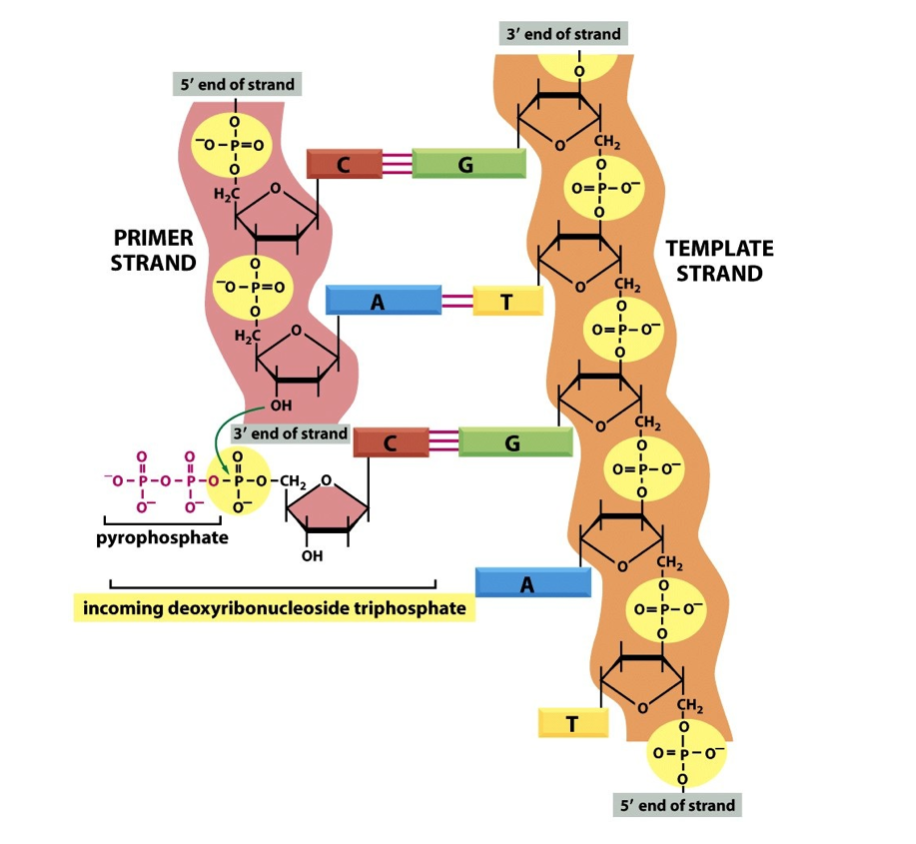
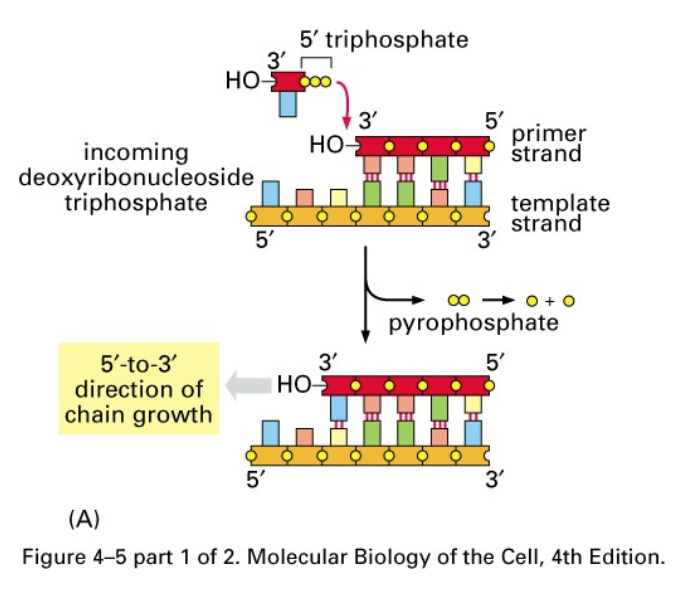
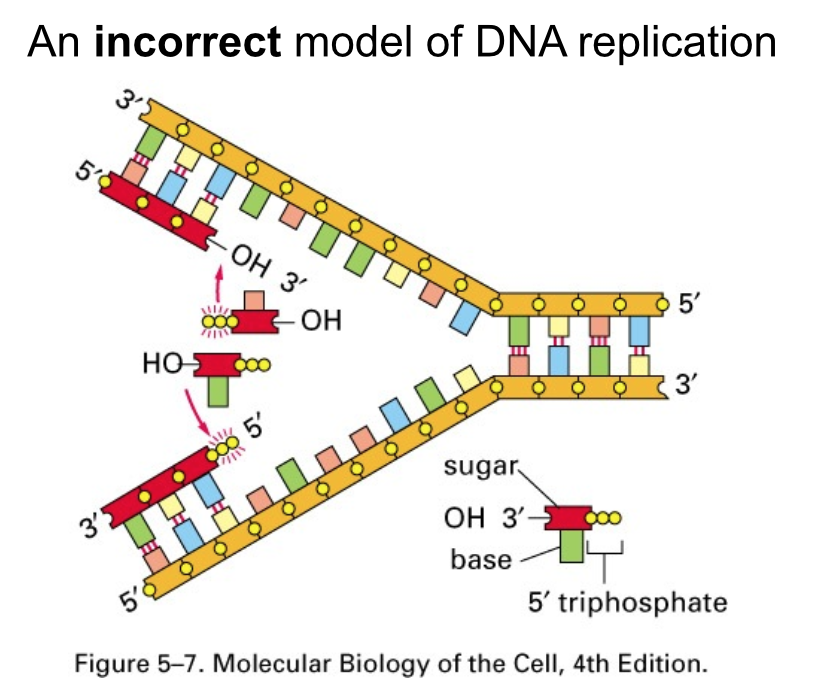
- DNA synthesis
- occurs in a 5’ -> 3’ direction
by breakage and formation
of phosphodiester bonds
- Strand orientation is antiparallel
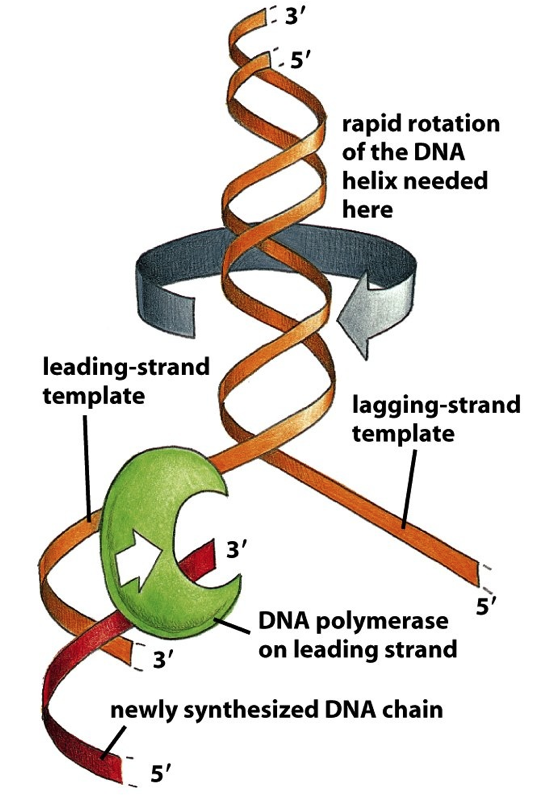
- Leading Strand and Lagging Strand synthesis are oriented
antiparallel to one another within the Replication Fork
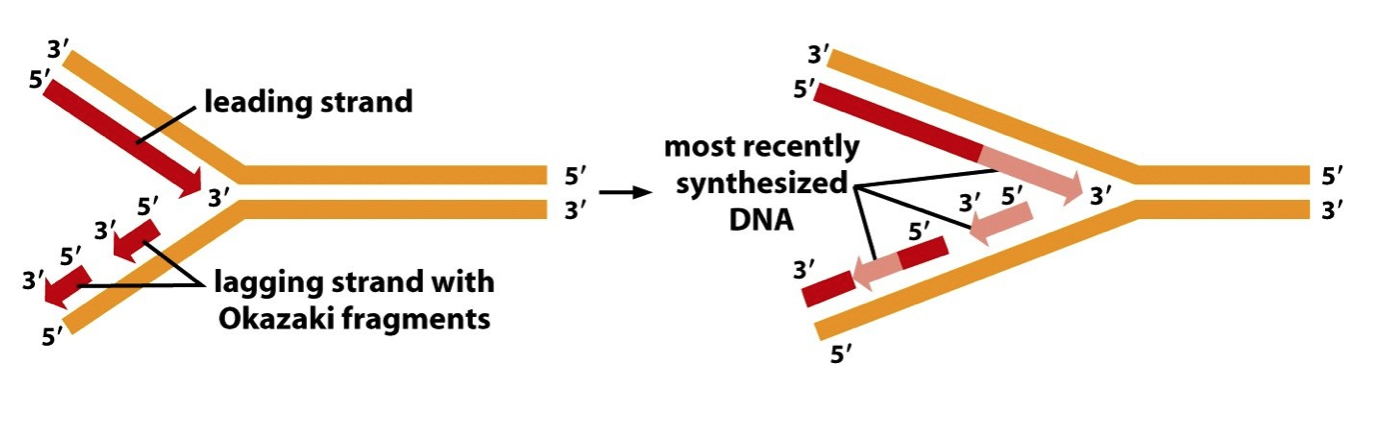
- Leading Strand synthesis is continuous and occurs 5’ -> 3’
- Lagging Strand synthesis is discontinuous and also occurs 5’ -> 3’
- Leading Strand synthesis is continuous and occurs 5’ -> 3’
- Leading Strand and Lagging Strand synthesis are oriented
antiparallel to one another within the Replication Fork
- Strand orientation is antiparallel
- effectively an irreversible reaction because it
is coupled to breakdown of PPi to 2Pi by
pyrophosphatase
- The free energy required for DNA
synthesis is provided by the breakage of
2 high energy phosphate bonds
- The free energy required for DNA
synthesis is provided by the breakage of
2 high energy phosphate bonds
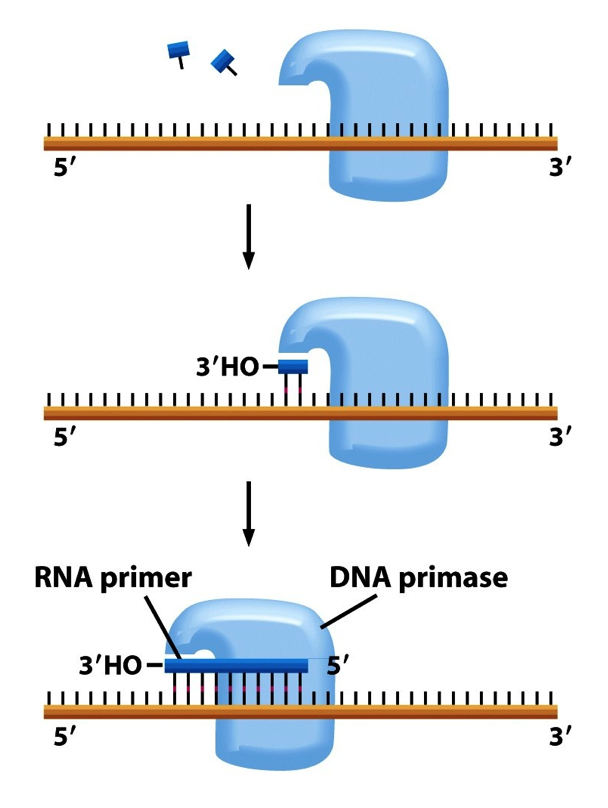
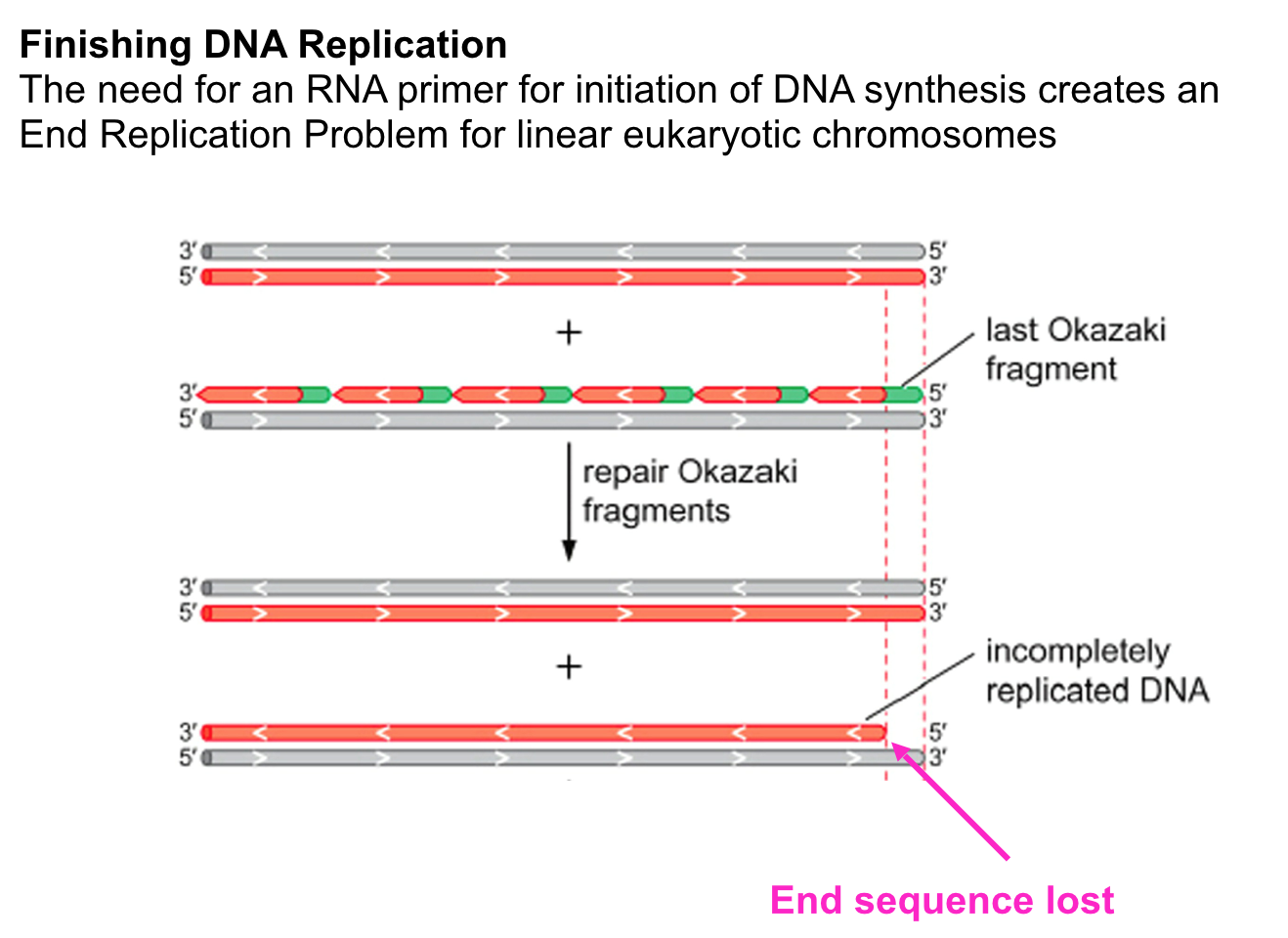
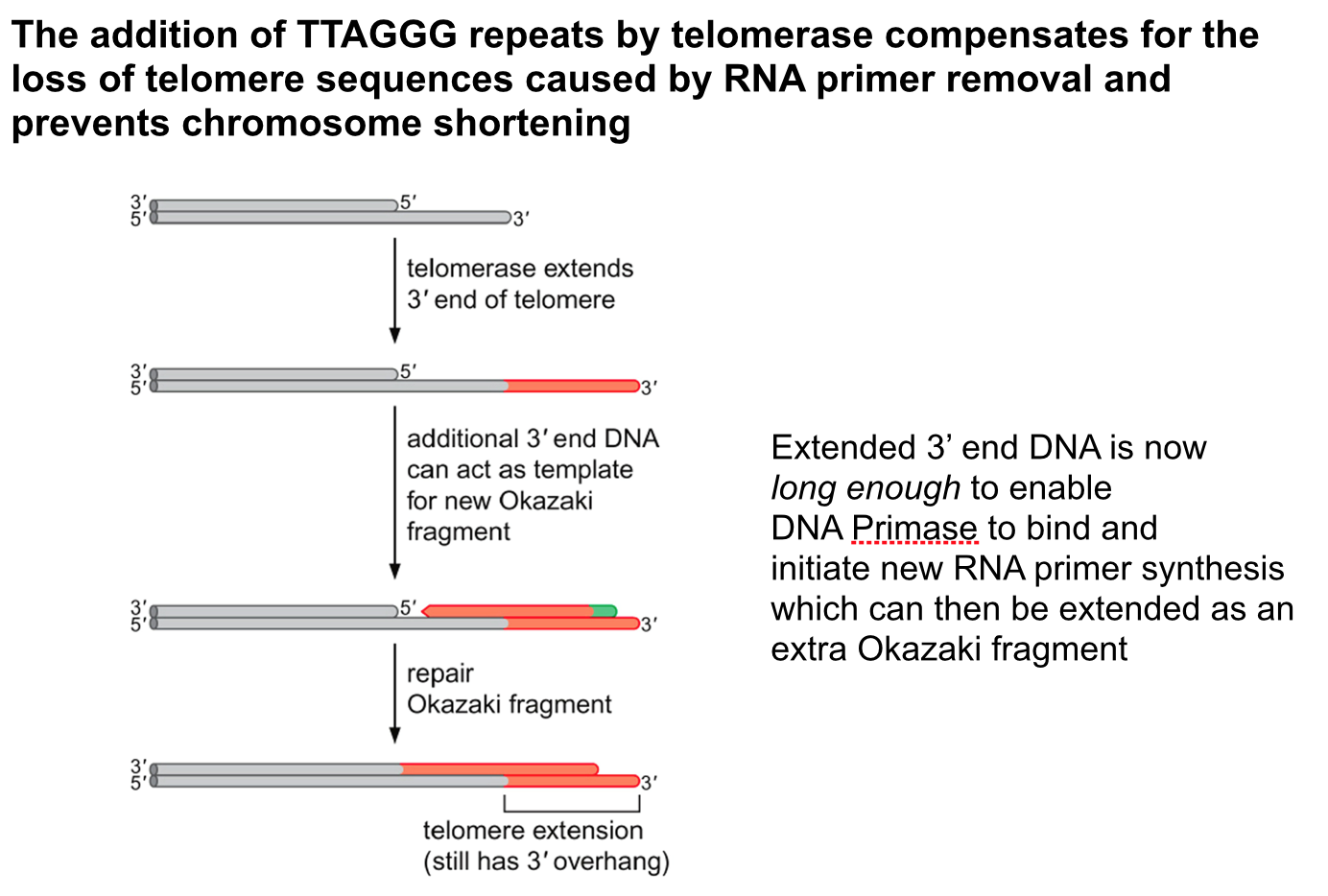
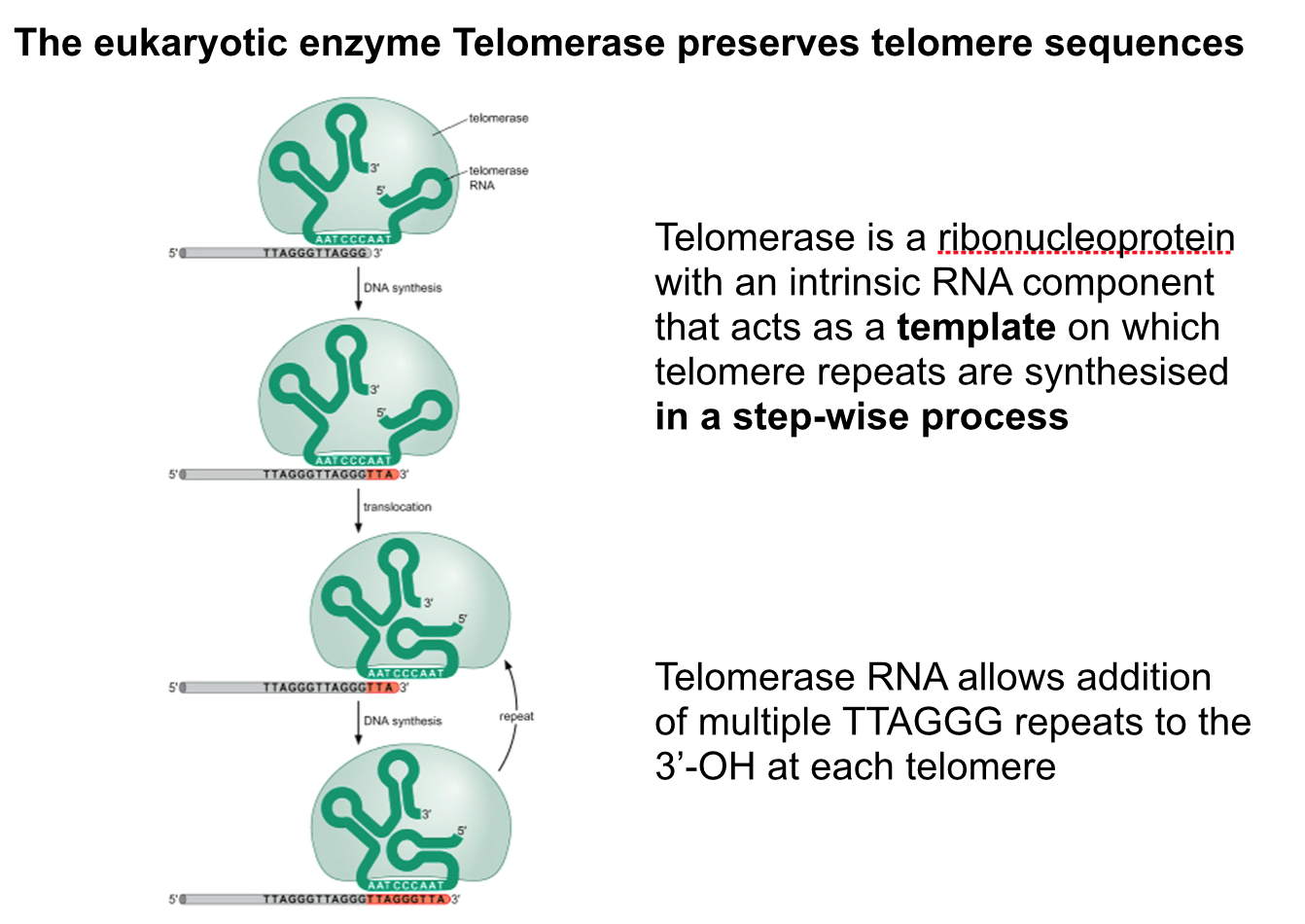
- All DNA synthesis is initiated by
extension of a short primer made
of RNA
- The RNA primer is
synthesised by DNA Primase
and only requires a DNA
template and NTPs
- A surprising role for
RNA in the initiation of
DNA Synthesis!
- A surprising role for
RNA in the initiation of
DNA Synthesis!
- The RNA primer is
synthesised by DNA Primase
and only requires a DNA
template and NTPs
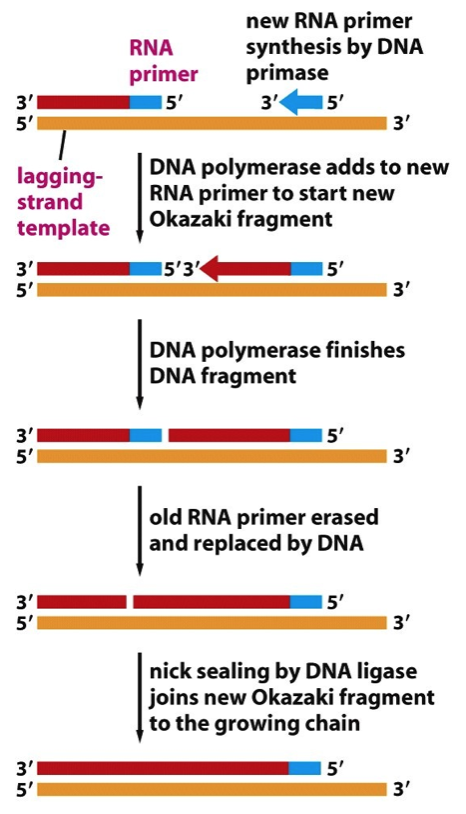
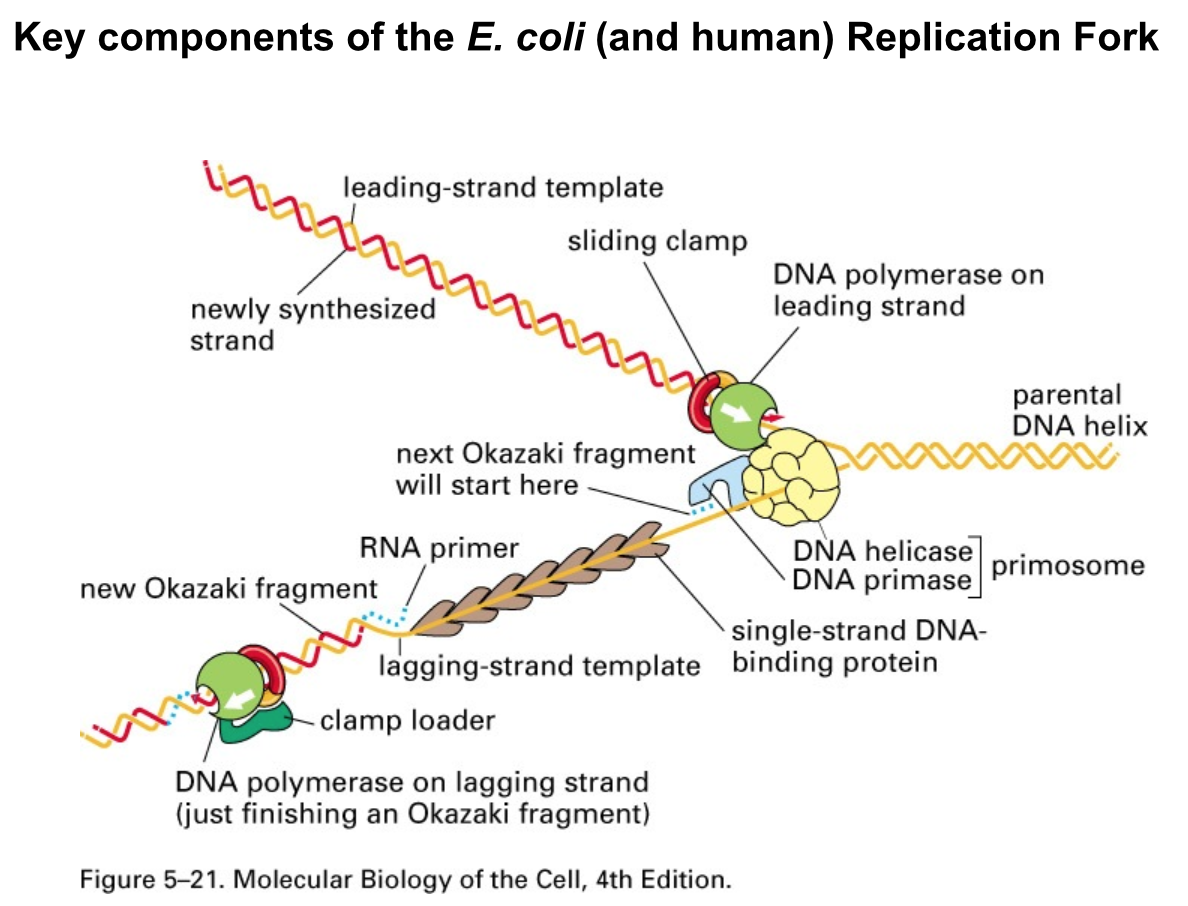
- Lagging Strand synthesis requires DNA
Primase, DNA Polymerase, Ribonuclease H
and DNA Ligase to convert Okazaki
fragments into a continuous strand of DNA
- DNA Primase –
makes RNA primer
- DNA Polymerase –
extends RNA primer
- Ribonuclease H –
removes RNA primer
- DNA Polymerase –
extends across gap
- DNA Ligase seals the nick
- DNA Ligase uses the energy of ATP hydrolysis to ligate newly
synthesised, adjacent DNA fragments in a two-step catalytic reaction
- The ligation process is rendered energetically
highly favourable by the conversion of PPi to 2Pi
by pyrophosphatase
- The ligation process is rendered energetically
highly favourable by the conversion of PPi to 2Pi
by pyrophosphatase
- DNA Ligase uses the energy of ATP hydrolysis to ligate newly
synthesised, adjacent DNA fragments in a two-step catalytic reaction
- DNA Primase –
makes RNA primer
- occurs in a 5’ -> 3’ direction
by breakage and formation
of phosphodiester bonds
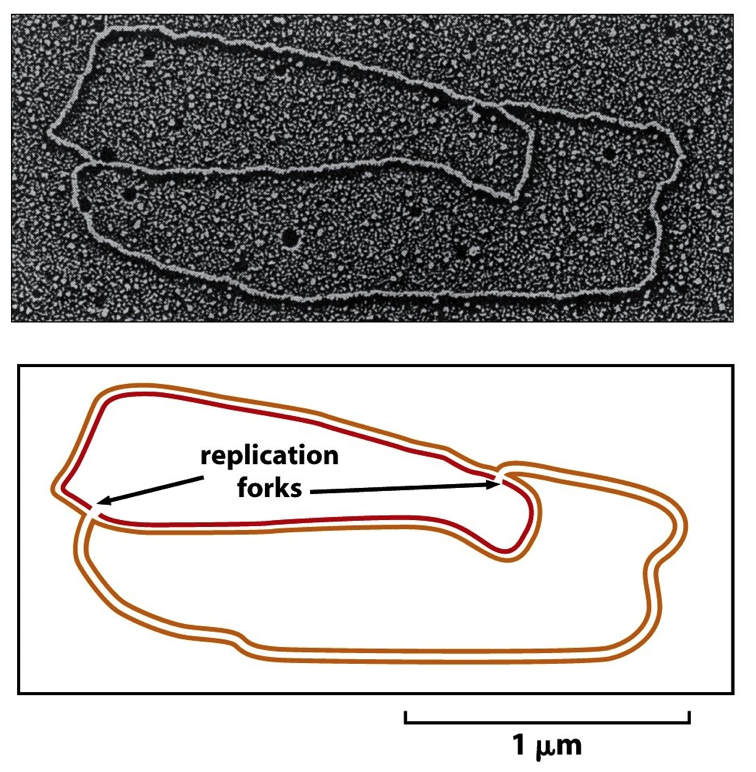
- DNA Replication in cells is bidirectional:
2 Replication Forks are created that
move in opposite directions
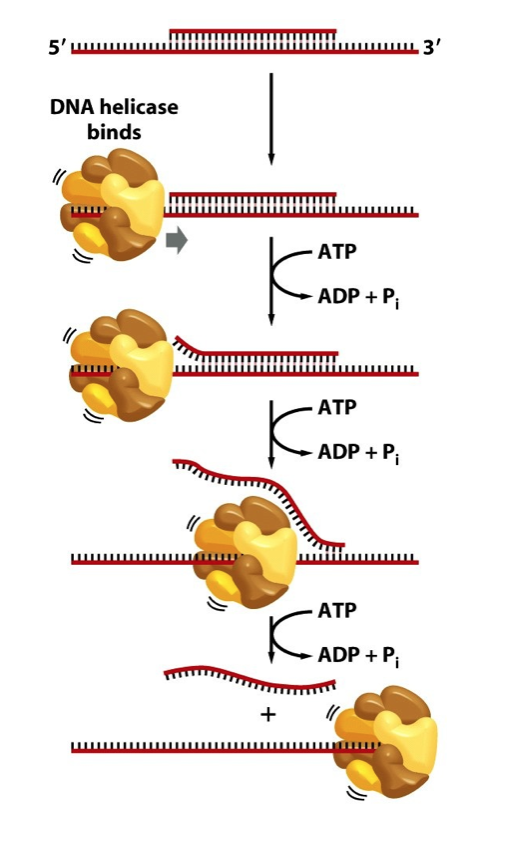
- DNA Helicase uses ATP to separate parental DNA strands at
the Replication Fork and move the Replication Fork forward
- A simple DNA strand separation assay reveals that DNA
Helicase activity is ATP-dependent and Magnesium-dependent
- rBLM protein is an ATP-dependent, Mg-dependent DNA Helicase that unwinds DNA
- Mutations in genes encoding DNA helicases cause human
diseases such as Werner Syndrome: a progeria (premature ageing)
- Werner Syndrome mutations are
autosomal recessive, occurring in RECQ
helicase gene WRN
- Werner Syndrome mutations are
autosomal recessive, occurring in RECQ
helicase gene WRN
- A simple DNA strand separation assay reveals that DNA
Helicase activity is ATP-dependent and Magnesium-dependent
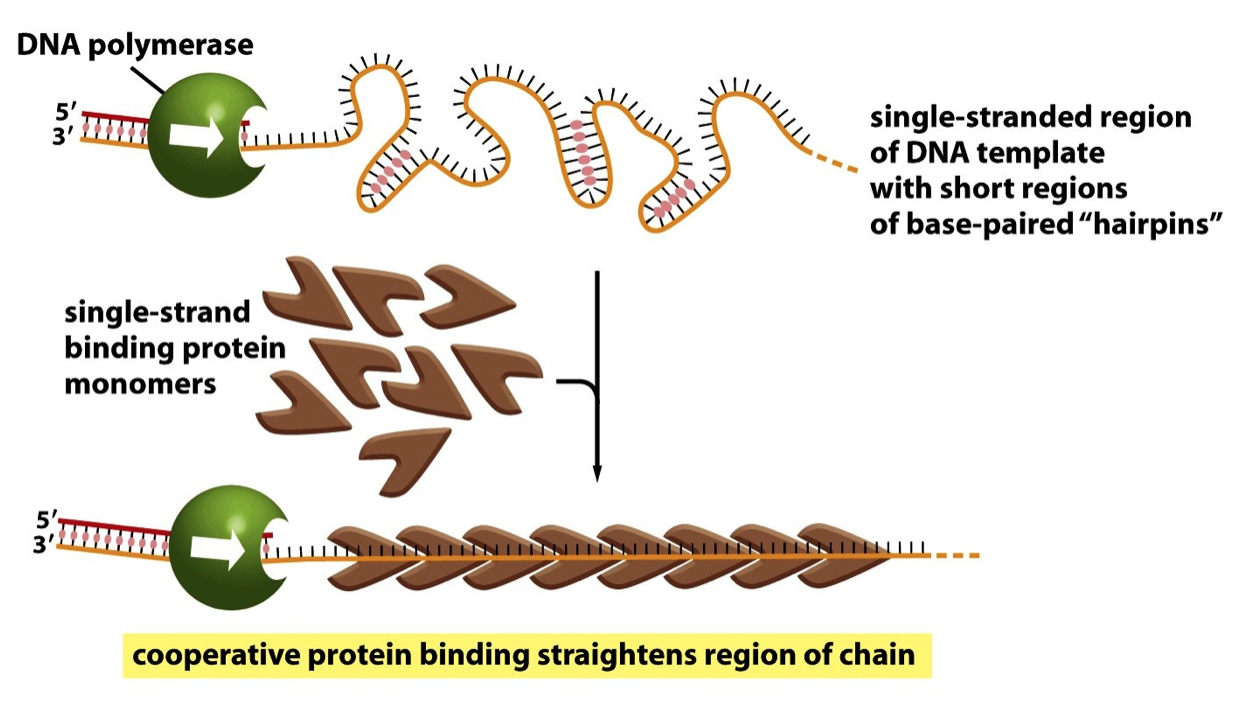
- Single-stranded
Binding Proteins
(SSBs)
- expose
single-stranded
DNA in the
replication fork
- making it available for
templating synthesis
of the new DNA strand
- and easing
replication fork
progression
- making it available for
templating synthesis
of the new DNA strand
- expose
single-stranded
DNA in the
replication fork
- DNA
topoisomerases
- prevent DNA from
becoming tangled during
DNA replication
- Unwinding of parental DNA
strands at the Replication
Fork introduces superhelical
tension into the DNA Helix.
- Tension is relaxed by DNA
Topisomerases, which nick and
reseal the backbone of the
parental helix
- Type I Topoisomerases nick and
reseal one of the 2 DNA strands, no
ATP required
- Type II Topoisomerases
nick and reseal both DNA
strands, ATP required (!)
- Type I Topoisomerases nick and
reseal one of the 2 DNA strands, no
ATP required
- Tension is relaxed by DNA
Topisomerases, which nick and
reseal the backbone of the
parental helix
- Unwinding of parental DNA
strands at the Replication
Fork introduces superhelical
tension into the DNA Helix.
- prevent DNA from
becoming tangled during
DNA replication
- DNA Helicase uses ATP to separate parental DNA strands at
the Replication Fork and move the Replication Fork forward
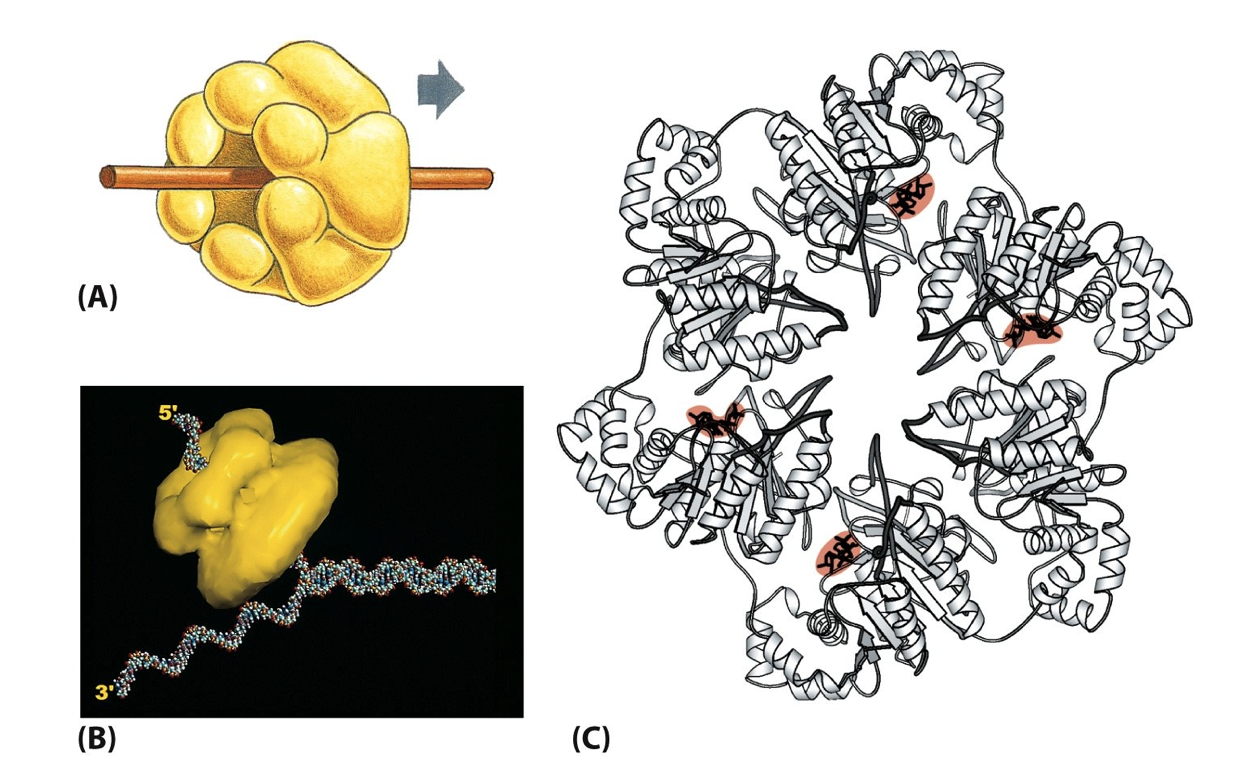
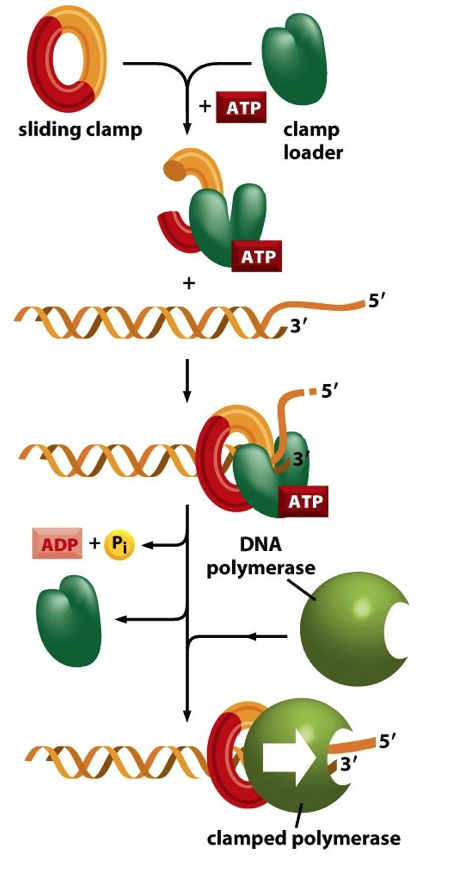
- The processivity of DNA Polymerases is greatly
enhanced by their association with a Sliding Clamp
- Processivity
- Once the first step of DNA synthesis has been accomplished,
interaction of enzyme with the Primer:Template junction is
maintained and addition of further nucleotides is very rapid
- Once the first step of DNA synthesis has been accomplished,
interaction of enzyme with the Primer:Template junction is
maintained and addition of further nucleotides is very rapid
- The Sliding Clamp (ATP-dependent!) is
positioned close to the Primer:Template
Junction by a Clamp Loader
- Sliding clamps encircle the DNA like a nut
on a bolt and help to move
DNA Polymerase forward
- Sliding clamps encircle the DNA like a nut
on a bolt and help to move
DNA Polymerase forward
- Processivity
- DNA replication requires the same
set of proteins in all organisms
- Helicase: disrupts base pairing in
dsDNA, enables replication fork
progress
- Single-stranded binding
proteins (SSBs): make
template an “easy read”
- Primase:
synthesises
RNA primer
- DNA Polymerase: extends primers annealed
to ssDNA template from 3’-OH end
- Sliding Clamp and Clamp Loader:
ensure processivity of DNA
Polymerase
- Ribonuclease H:
removes RNA primer
- DNA Ligase: ligates adjacent
single-stranded DNA fragments
- Topoisomerases:
break and rejoin
phosphodiester bonds
in DNA backbone,
relax supercoils
generated by Helicase
- Helicase: disrupts base pairing in
dsDNA, enables replication fork
progress
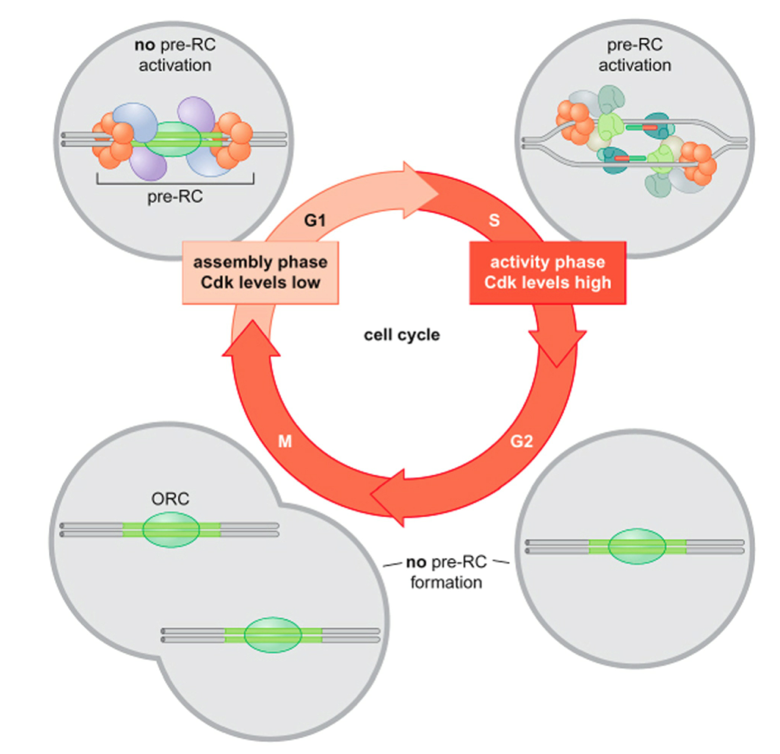
- Control of DNA Replication:
Initiation at replication origin
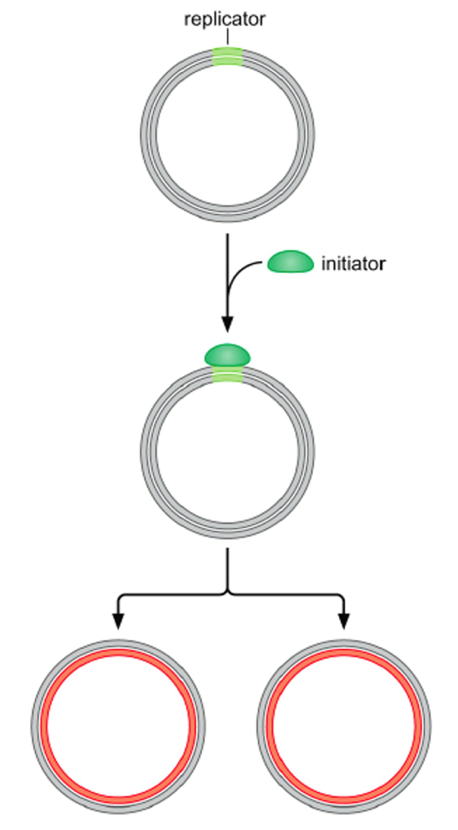
- Replicators - direct
the initiation of DNA
replication by
recruiting Replication
Initiator proteins
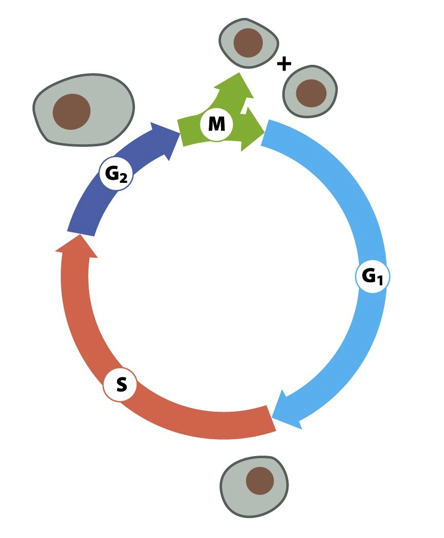
- Initiation of DNA replication in
eukaryotes is biphasic:
- Replicator Selection -
formation of a pre-Replicative
Complex - occurs in G1 phase
- Eukaryotic Replicator Selection occurs in
G1 and leads to the formation of a
Pre-Replicative Complex (pre-RC)
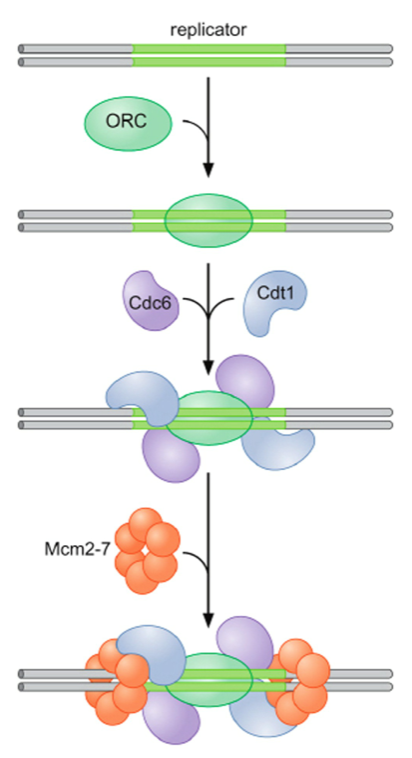
- Origin Recognition
Complex Binds to
Replicator sequence
- Helicase loading
proteins Cdc6 and Cdt1
bind to ORC
- The Helicase Mcm2-7
binds to complete
formation of pre-RC
- The Helicase Mcm2-7
binds to complete
formation of pre-RC
- Helicase loading
proteins Cdc6 and Cdt1
bind to ORC
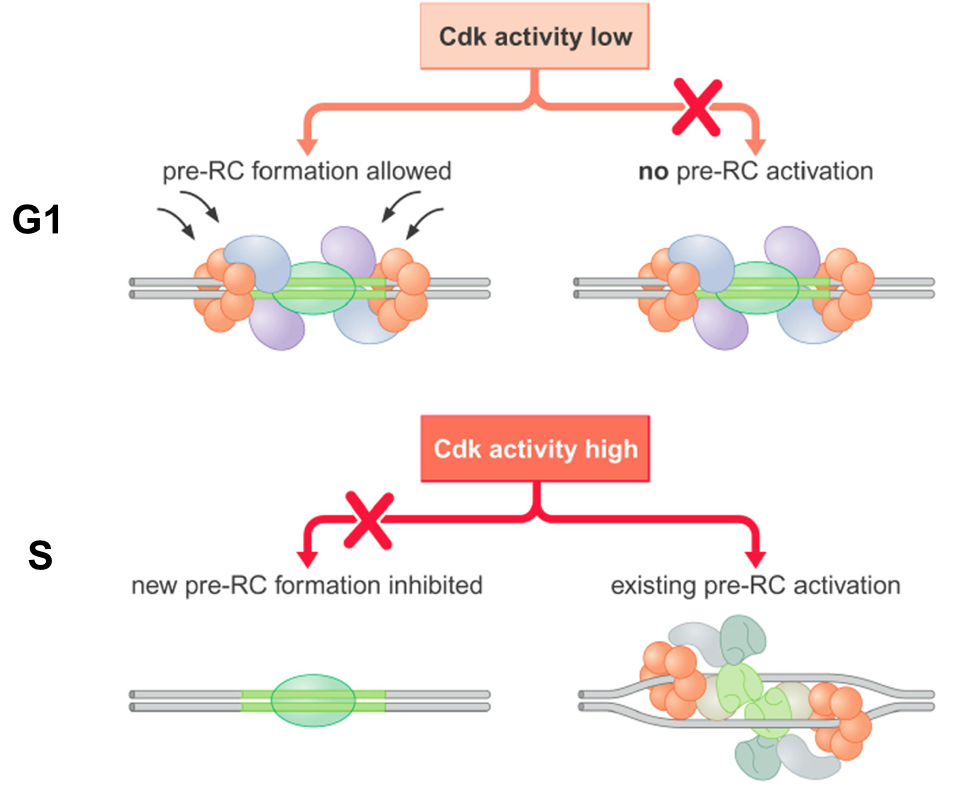
- High levels of Cyclin-dependent kinase
(Cdk) activity in S-phase activates existing
pre-RC but prevents formation of new
pre-RCs
- Close relationships between pre-RC function, Cdk levels
and cell cycle ensures that chromosomes are replicated
exactly once per cell cycle
- Origin Recognition
Complex Binds to
Replicator sequence
- Eukaryotic Replicator Selection occurs in
G1 and leads to the formation of a
Pre-Replicative Complex (pre-RC)
- Origin Activation - unwinding of
DNA and recruitment of DNA
Polymerase - occurs in S phase
- Temporal separation of these
2 events ensures that each
chromosome is only replicated
exactly once per cell cycle
- Replicator Selection -
formation of a pre-Replicative
Complex - occurs in G1 phase
- Replicators - direct
the initiation of DNA
replication by
recruiting Replication
Initiator proteins
Media attachments
{kind=link}
{kind=link}
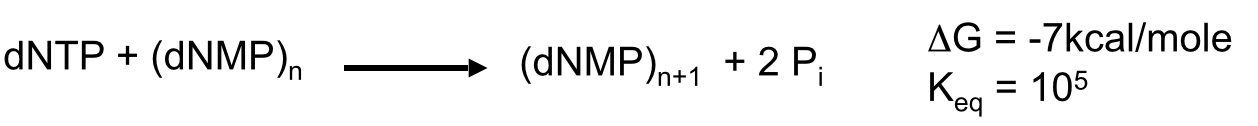{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
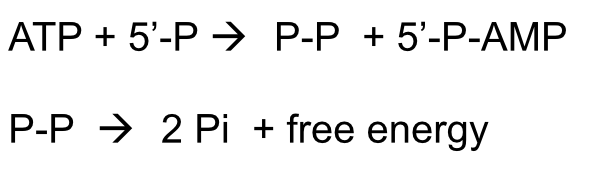{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Want to create your own Mind Maps for free with GoConqr? Learn more.