Description
|
|
Created by Cher Bachar
over 12 years ago
|
|
Page 1
{kind=link}
{kind=link}
{kind=link}
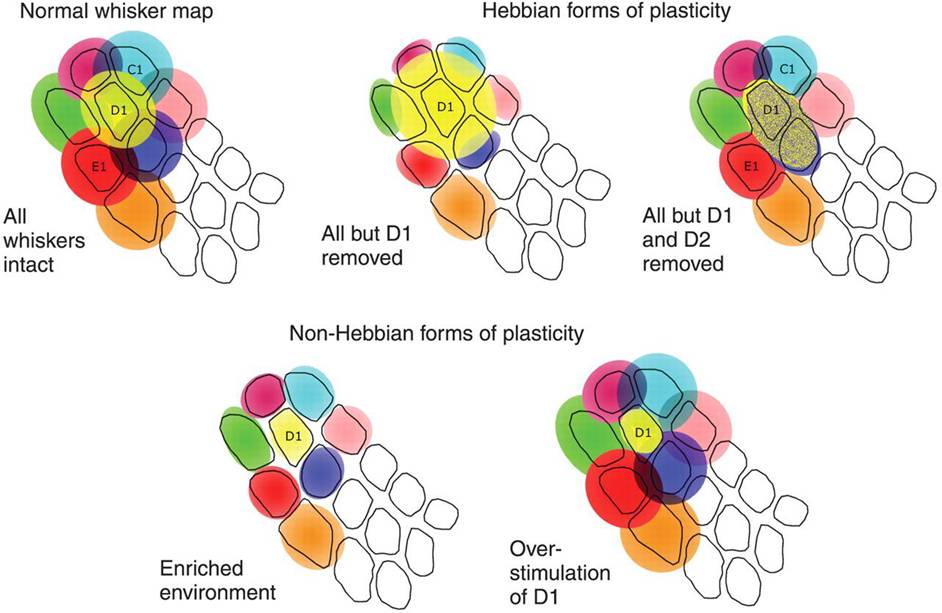
Fig: When all whiskers are intact the receptive fields stay their relevant space, however when for example all but 1 whisker are removed (D1) the receptive field grows to try to compensate to the loss in sensory input, however the resolution would decrease massively. See fig for other situations.
{kind=link}
{kind=link}
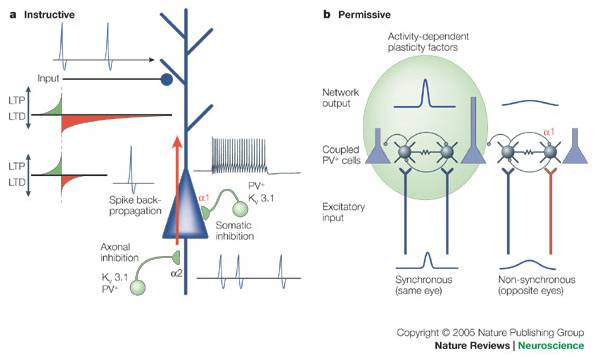
a | Somatic inhibition by fast-spiking parvalbumin-positive (PV+) cells (green) expressing the potassium channel Kv3.1 is mediated by the GABAA (γ-aminobutyric acid type A) α1-subunit, whereas axonal inhibition is mediated by the GABAA α2-subunit. Somatic inhibition is ideally situated for suppression, or 'editing' of unwanted spikes, preventing them from back-propagating through the cell body into the dendritic tree (red arrow). This is an 'instructive' model, as individual action potentials can produce long-term potentiation (LTP) or depression (LTD) based on precise spike-timing dependent windows at individual synapses of coincident pre- and postsynaptic activity123, 124. Notably, failure to regulate excess spiking at the axon initial segment can still be differentiated by fast-spiking inhibition at the cell body. b | Gap junctional coupling endows networks of parvalbumin-positive (PV+) interneurons with the ability to detect synchronous input128, 129, 130. Even a slight jitter in input timing (for example, between eyes) dampens network activity through reciprocal GABA-mediated contacts (enriched with GABAA-receptor α1-subunits131). Only synchronous open-eye input will produce maximal, activity-dependent release or uptake of 'permissive' factors for neurite growth (for example, tissue-type plasminogen activator78, 86 and brain-derived neurotrophic factor). Competition is determined extracellularly.
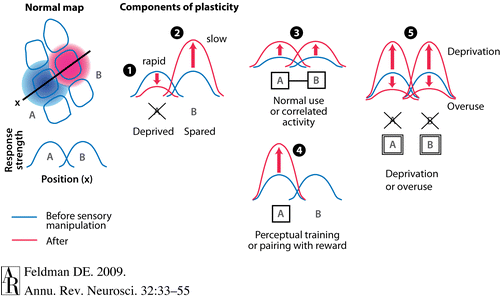
components of cortical plasticity
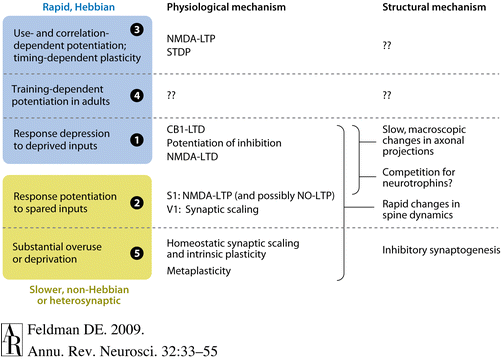
mechanisms
barrel cortex plasticity
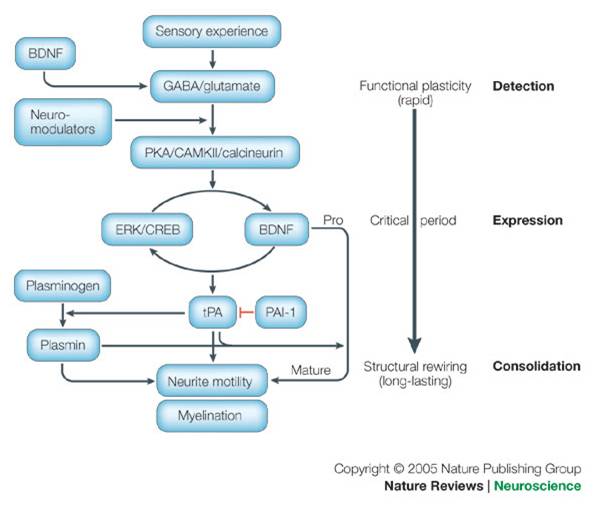
molecular mechanisms of visual plasticity/ circuit plasticit
Want to create your own Notes for free with GoConqr? Learn more.