28633477
Descrição
Mapa Mental por Ximena Porras, atualizado more than 1 year ago
|
|
Criado por Ximena Porras
quase 4 anos atrás
|
|
Percepción del Color
- La percepción del color en el ser humano se produce mediante la recepción de la luz por parte del ojo y su proyección en su parte
posterior, la retina, que contiene células especializadas para detectar las distintas longitudes de onda presentes. Estas células
generan impulsos nerviosos que, a través del nervio óptico, llegan a la corteza visual, la parte del cerebro encargada del
procesamiento e interpretación de los estímulos visuales. Analizaremos en mayor detalle estas etapas.
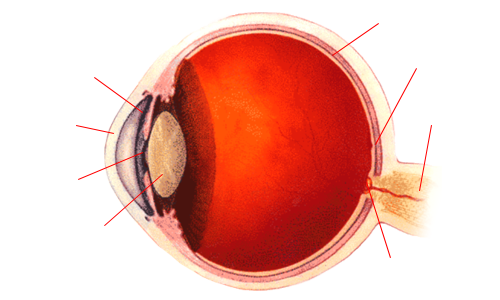
- EL OJO
- Desde el punto de vista óptico, el ojo humano funciona como una cámara oscura
con una abertura para el paso de la luz (la pupila), un lente para enfocarla (el
cristalino) y una superficie donde la imagen se proyecta y detecta (la retina). Esta
última es la que participa en el proceso de la percepción del color.
Anotações:
- Diagrama esquemático del ojo humano, visto desde arriba (ojo derecho), con indicación de las partes relevantes a la visión de color.
- Los impulsos nerviosos se conducen al cerebro mediante el nervio óptico;
el punto de la retina donde estos confluyen no tiene capacidad de detectar
luz: es el punto ciego.
- Desde el punto de vista óptico, el ojo humano funciona como una cámara oscura
con una abertura para el paso de la luz (la pupila), un lente para enfocarla (el
cristalino) y una superficie donde la imagen se proyecta y detecta (la retina). Esta
última es la que participa en el proceso de la percepción del color.
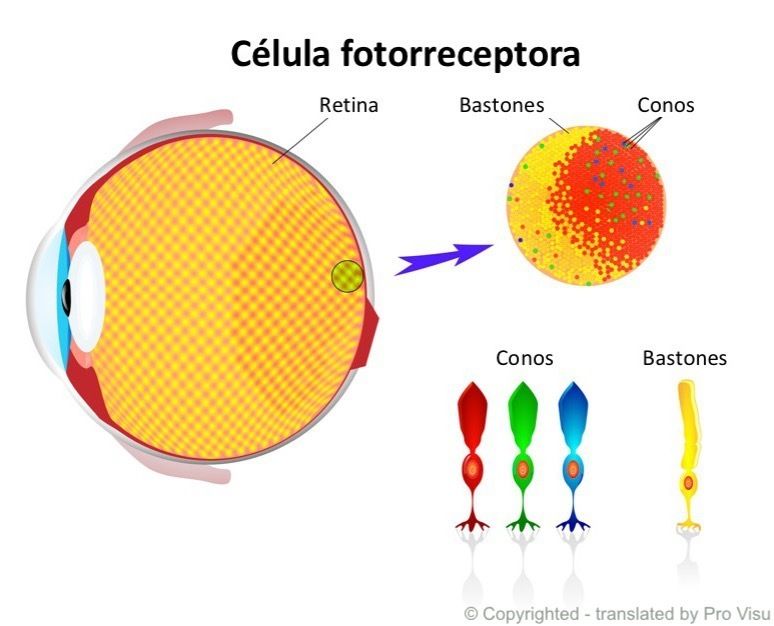
- LA RETINA
- La retina consta de la pared posterior del ojo, y allí es donde se proyecta la imagen.
La homogeneidad de su superficie sólo se interrumpe en dos lugares: el punto ciego,
que es el lugar de donde parte el nervio óptico hacia el cerebro, y la fóvea, una
pequeña depresión ubicada justo en el eje óptico, en un lugar aproximadamente
opuesto a la pupila.
- Distribuidas en toda su superficie, la retina contiene células de
dos tipos: los bastones y los conos.
- LOS BASTONES
- Los bastones son sensibles sólo a bajas intensidades de luz y prácticamente dejan de ser útiles a intensidades medias a elevadas. Sólo detectan
cantidad de luz y no tienen discriminación de color, aunque no responden igualmente a cualquier longitud de onda: son más sensibles en el rango
azul-verde (aproximadamente 500 nm) y prácticamente insensibles al rojo (640 nm o más). Hay en la retina entre 90 y 100 millones en promedio y
son mucho más numerosos que los conos (aproximadamente 13:1). Tienden a concentrarse más hacia la parte externa de la retina y por lo tanto
son los que dominan nuestra visión periférica. Son responsables de la visión escotópica, la que opera en condiciones de penumbra o de visión
nocturna, y son alrededor de 100 veces más sensibles que los conos. Se los representa con la letra R (por rods, bastones).
- Los bastones son sensibles sólo a bajas intensidades de luz y prácticamente dejan de ser útiles a intensidades medias a elevadas. Sólo detectan
cantidad de luz y no tienen discriminación de color, aunque no responden igualmente a cualquier longitud de onda: son más sensibles en el rango
azul-verde (aproximadamente 500 nm) y prácticamente insensibles al rojo (640 nm o más). Hay en la retina entre 90 y 100 millones en promedio y
son mucho más numerosos que los conos (aproximadamente 13:1). Tienden a concentrarse más hacia la parte externa de la retina y por lo tanto
son los que dominan nuestra visión periférica. Son responsables de la visión escotópica, la que opera en condiciones de penumbra o de visión
nocturna, y son alrededor de 100 veces más sensibles que los conos. Se los representa con la letra R (por rods, bastones).
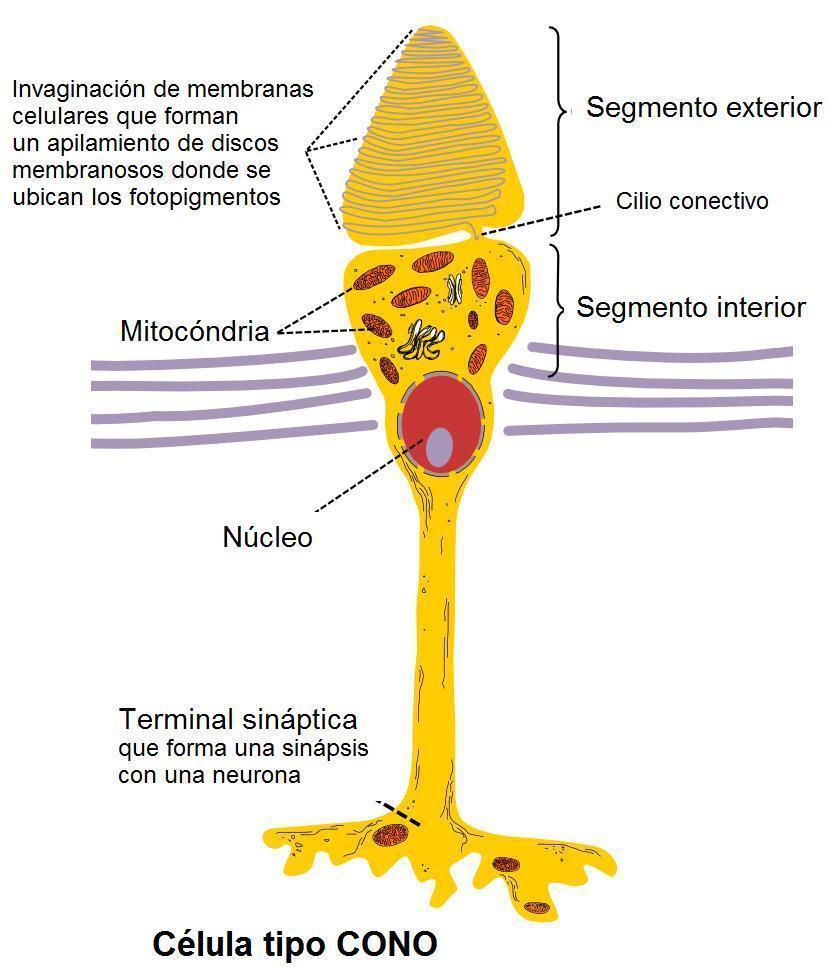
- LOS CONOS
- Los conos son estructuralmente similares a los bastones, pero requieren mucha mayor intensidad de luz para volverse activos. Operan en
condiciones de luz diurna y son responsables de la visión fotópica, donde ocurre la auténtica percepción del color. Son menos numerosos
(alrededor de 7 millones) y tienden a concentrarse en la fóvea. A diferencia de los bastones, existen tres tipos de conos sensibles a una parte
particular del espectro visible. Individualmente, cada uno responde a la cantidad de luz que se concentra en el rango de longitudes de onda
correspondiente; la discriminación de color aparece por la diferencia relativa en la excitación de cada uno. Por ejemplo, dado un espectro
particular, el color que se percibe del mismo dependerá de cuanta energía ese espectro tiene en el rango de sensibilidad de cada cono. Por lo
tanto, dos espectros luminosos distintos que produzcan en la retina los mismos estímulos serán percibidos necesariamente como el mismo
color, dado que los impulsos
Anotações:
- La visión humana del color se denomina tricromática debido a que el ojo consta de tres sensores independientes (los conos L, M y S) que operan en zonas diferentes del espectro visible. Esto explica por qué los sistemas que cuantifican el color requieren siempre tres parámetros.
- Los conos se denominan en función del rango de longitudes de
onda en el que operan:
- Conos S (por short, corto)
- Trabajan en el rango de longitudes de onda más cortas (zona
azul del espectro), con un máximo de sensibilidad en 420 nm.
- Trabajan en el rango de longitudes de onda más cortas (zona
azul del espectro), con un máximo de sensibilidad en 420 nm.
- Conos M (por medium)
- Son sensibles a la zona de los verdes, con un
máximo en los 530 nm.
- Son sensibles a la zona de los verdes, con un
máximo en los 530 nm.
- Conos L (por large, grande)
- Operan en longitudes de onda algo más largas que los M
(zona del verde-amarillo), con un pico de sensibilidad en
los 560 nm.
- Operan en longitudes de onda algo más largas que los M
(zona del verde-amarillo), con un pico de sensibilidad en
los 560 nm.
- Conos S (por short, corto)
- Los conos se denominan en función del rango de longitudes de
onda en el que operan:
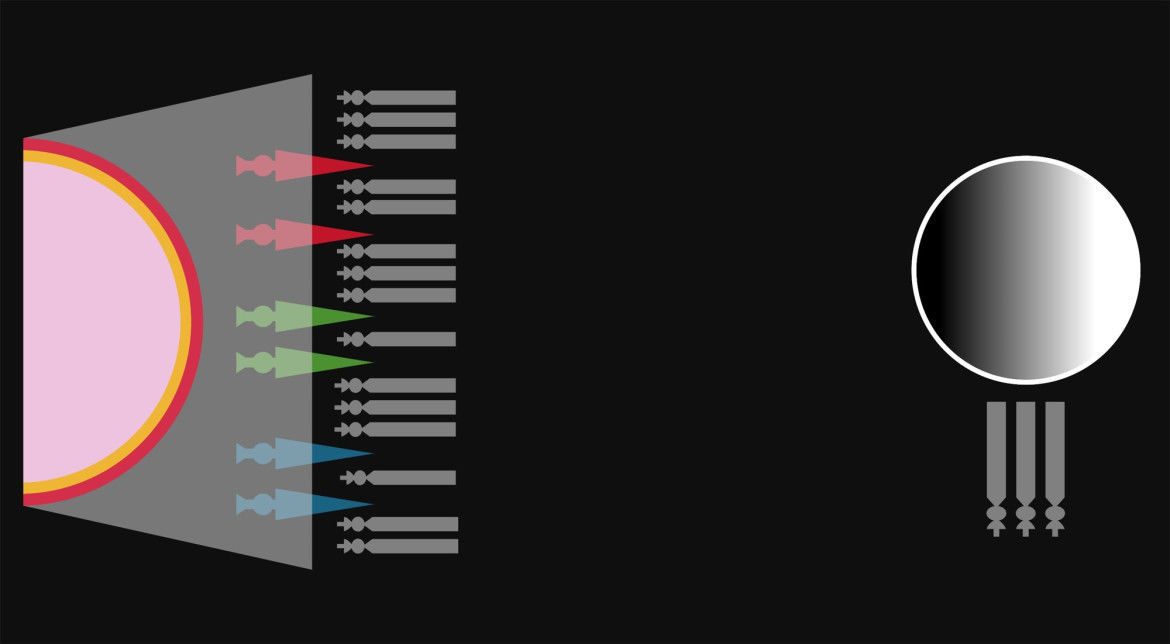
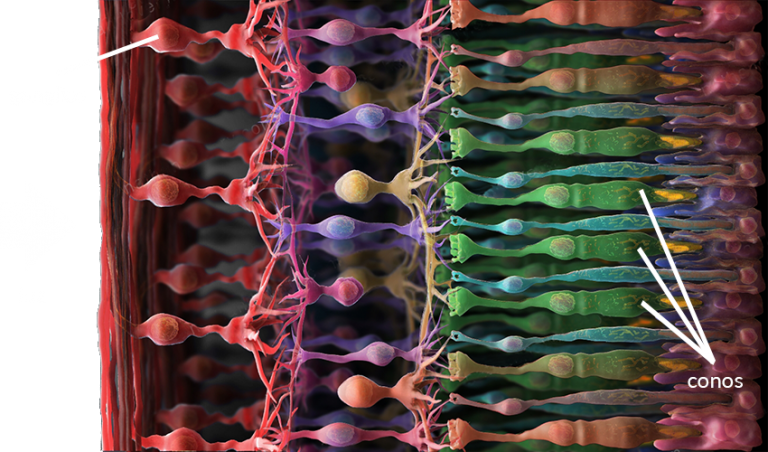
- Los conos no transmiten los impulsos nerviosos directamente al cerebro.
Ciertas células especializadas llamadas ganglios toman esos impulsos de
grupos de conos, procesan esas señales y entregan el resultado al nervio
óptico.
Anotações:
- Corte transversal de la retina. En este diagrama, la luz incide desde la izquierda e incide en los conos ubicados sobre la pared posterior. Los ganglios integran (suman) la respuesta de varios conos y envían el resultado a un nervio.
- Las señales nerviosas procesadas por los ganglios resultan de la comparación de ciertas
combinaciones de los impulsos de los conos L, M y S. Por ejemplo, algunos ganglios
responden a la suma de los impulsos de todos los conos y son los responsables de generar la
escala negro-blanco, ya que obtiene el brillo absoluto del color percibido. El proceso explica
la imposibilidad fisiológica de percibir ciertos tonos «imposibles», como un color rojo-verde,
y justifica la llamada teoría de los procesos por oposición:
- La teoría de los procesos por oposición se desarrolló para explicar la inexistencia de ciertas
combinaciones de colores (blanco-negro, rojo-verde, azul-amarillo) y otros fenómenos como
el de las imágenes latentes (las percibidas luego de una larga exposición a un color
determinado), y apuntan a cómo se genera la percepción de color, más que la forma en la
que se detecta.
- La teoría de los procesos por oposición se desarrolló para explicar la inexistencia de ciertas
combinaciones de colores (blanco-negro, rojo-verde, azul-amarillo) y otros fenómenos como
el de las imágenes latentes (las percibidas luego de una larga exposición a un color
determinado), y apuntan a cómo se genera la percepción de color, más que la forma en la
que se detecta.
- Los conos son estructuralmente similares a los bastones, pero requieren mucha mayor intensidad de luz para volverse activos. Operan en
condiciones de luz diurna y son responsables de la visión fotópica, donde ocurre la auténtica percepción del color. Son menos numerosos
(alrededor de 7 millones) y tienden a concentrarse en la fóvea. A diferencia de los bastones, existen tres tipos de conos sensibles a una parte
particular del espectro visible. Individualmente, cada uno responde a la cantidad de luz que se concentra en el rango de longitudes de onda
correspondiente; la discriminación de color aparece por la diferencia relativa en la excitación de cada uno. Por ejemplo, dado un espectro
particular, el color que se percibe del mismo dependerá de cuanta energía ese espectro tiene en el rango de sensibilidad de cada cono. Por lo
tanto, dos espectros luminosos distintos que produzcan en la retina los mismos estímulos serán percibidos necesariamente como el mismo
color, dado que los impulsos
- LOS BASTONES
- Distribuidas en toda su superficie, la retina contiene células de
dos tipos: los bastones y los conos.
- La retina consta de la pared posterior del ojo, y allí es donde se proyecta la imagen.
La homogeneidad de su superficie sólo se interrumpe en dos lugares: el punto ciego,
que es el lugar de donde parte el nervio óptico hacia el cerebro, y la fóvea, una
pequeña depresión ubicada justo en el eje óptico, en un lugar aproximadamente
opuesto a la pupila.
- TEORÍAS
Anotações:
- Analizado en conjunto, este análisis muestra por qué tanto la teoría tricromática como la teoría del proceso por oposición explican satisfactoriamente la visión de color: en esencia, ambos describen partes separadas el proceso: *La teoría tricromática explica la captura de la información cromática de la luz incidente. *La teoría del proceso por oposición explica el proceso de comparación intermedio al que se somete la información captada por los conos.
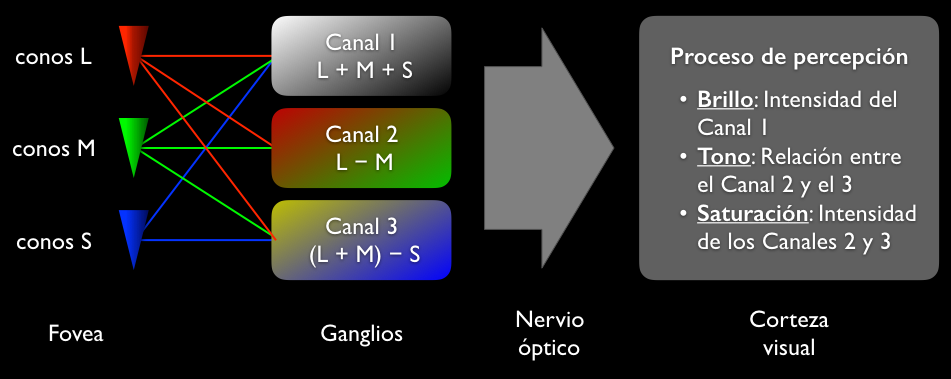
- La información así procesada es conducida por el nervio óptico a la corteza visual, donde tiene lugar la auténtica percepción del color, de modo que: -La percepción del brillo del color proviene del canal 1. -Lo que identificamos como matiz o tono es una relación entre los canales 2 y 3. Como estos dos canales son independientes, esta relación ocupa una posición en un plano y justifica asignar al tono un ángulo; de allí la posibilidad de representar el matiz mediante un círculo cromático.-Lo que percibimos como saturación es la intensidad de ambos canales 2 y 3, es decir, que tan inclinados están hacia uno de sus extremos; como un canal balanceado equivale a una percepción acromática, esta cantidad indica que tan lejos esta el color percibido de esa percepción.
- TEORÍA DE PROCESOS DEL COLOR POR OPOSCICIÓN
- Fue propuesta por Ewald Heringen 1872, el principio biológico de la teoría del
proceso por oposición descansa en la existencia de los ganglios, que en un
primer análisis simplificado se pueden diferenciar tres grupos:
Anotações:
- Diagrama del proceso de visión del color en el ser humano.
- Los que procesan la intensidad absoluta de la luz recibida, a partir de la suma de la respuesta de
todos los conos: son los responsables del eje blanco-negro (canal 1).
- Los que procesan la diferencia de excitación entre los conos L y M: son los responsables de
definir el eje rojo-verde del color percibido (canal 2).
- Los que lo hacen entre una suma de los L y M y los conos S, y son los que
definen el eje azul-amarillo (canal 3).
- Fue propuesta por Ewald Heringen 1872, el principio biológico de la teoría del
proceso por oposición descansa en la existencia de los ganglios, que en un
primer análisis simplificado se pueden diferenciar tres grupos:
- TEORÍA TRICROMÁTICA
- Propuesta por Thomas Young en 1802 y reelaborada por Herman Helmholtz
en 1852. Sostiene que la percepción del color es el resultado de la interacción
de tres mecanismos receptores que poseen diferentes sensibilidades
espectrales, es decir, que son sensibles a distintas longitudes de onda.
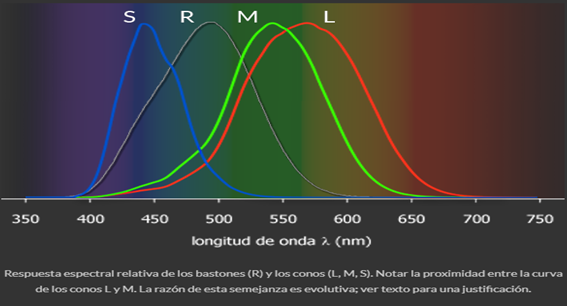
- SENSIBILIDAD ESPECTRAL DEL OJO HUMANO
- En el siguiente gráfico se observan las curvas de sensibilidad relativa de
bastones (R) y conos (L, M y S). Notar que estas curvas sólo indican el
rango de longitudes de onda a las que son sensibles y la posición en la
que presentan su máxima respuesta.
- Estas curvas nos permiten precisar dos nociones:
- El ojo no es sensible separadamente al rojo, verde y azul como es de creencia
común. Si bien el modelo RGB es una consecuencia de la manera en que
opera la visión humana, no existe una relación uno a uno con la respuesta
espectral de los conos.
- Los conos L y M tienen respuestas muy cercanas; la razón descansa en el proceso evolutivo que tuvo lugar en los mamíferos primitivos (que
sólo contaban con conos S y M) y que desembocó en los simios superiores y el hombre. Se piensa que una alteración genética de los conos
M produjo la creación de conos L con una sensibilidad espectral ligeramente desplazada al rojo, lo que pudo haber sido una ventaja
evolutiva importante para detectar el grado de maduración de vegetales y frutos, imprescindibles en la alimentación de los primeros
simios. La alteración de este gen puede dar lugar a la falta de desarrollo de los conos L o M, provocando alguna variante de daltonismo.
Como el gen responsable de este cambio se encuentra en el cromosoma X, del cual los hombres (que son XY) sólo poseen una copia, las
alteraciones en la visión de color son mucho más frecuentes que en las mujeres (que al poseer dos cromosomas X tienen mucha menor
posibilidad de presentar la alteración en ambos al mismo tiempo)
- El ojo no es sensible separadamente al rojo, verde y azul como es de creencia
común. Si bien el modelo RGB es una consecuencia de la manera en que
opera la visión humana, no existe una relación uno a uno con la respuesta
espectral de los conos.
- Estas curvas nos permiten precisar dos nociones:
- En el siguiente gráfico se observan las curvas de sensibilidad relativa de
bastones (R) y conos (L, M y S). Notar que estas curvas sólo indican el
rango de longitudes de onda a las que son sensibles y la posición en la
que presentan su máxima respuesta.
- SENSIBILIDAD ESPECTRAL DEL OJO HUMANO
- Propuesta por Thomas Young en 1802 y reelaborada por Herman Helmholtz
en 1852. Sostiene que la percepción del color es el resultado de la interacción
de tres mecanismos receptores que poseen diferentes sensibilidades
espectrales, es decir, que son sensibles a distintas longitudes de onda.
- TEORÍA DE LA LUZ
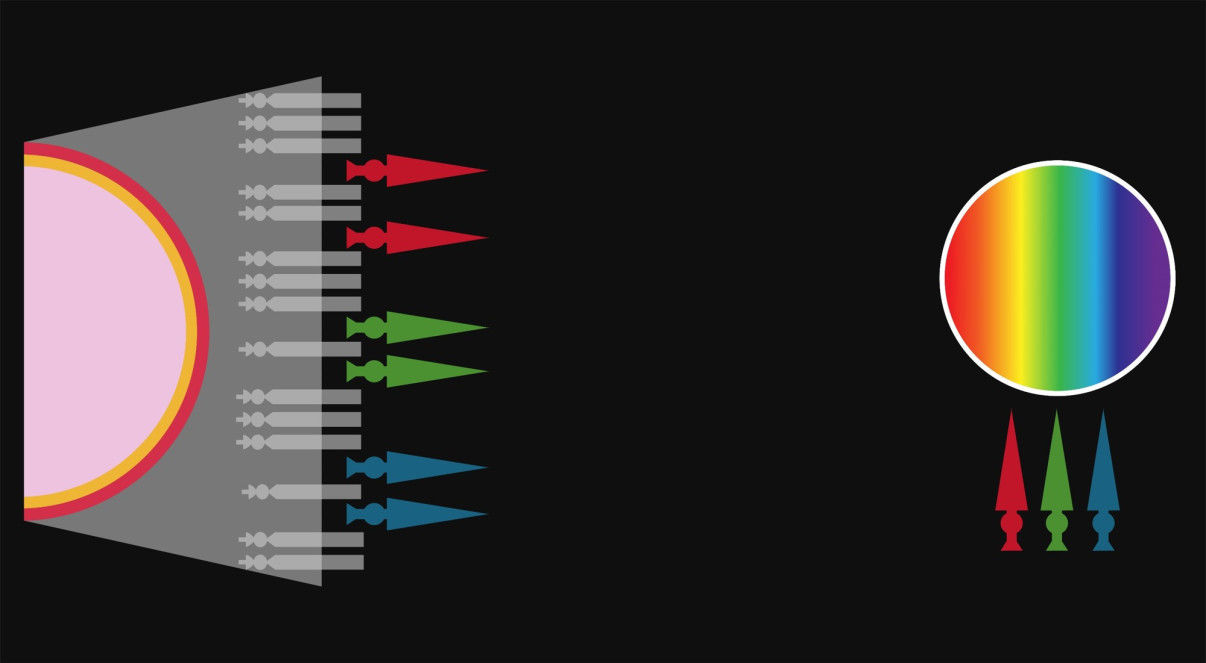
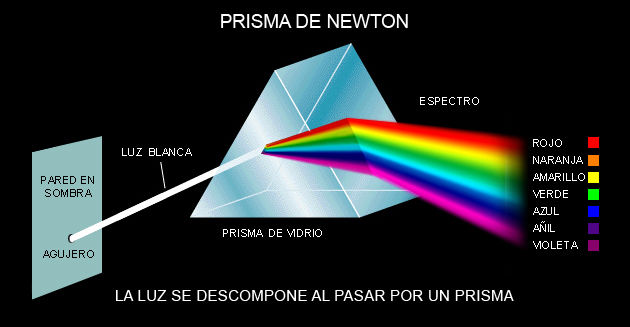
- Entre 1670 y 1672, Sir Isaac Newton demostró que la luz blanca estaba formada por una
banda de colores: rojo, naranja, amarillo, verde, azul, añil/índigo y violeta. Lo descubrió
mediante la proyección de esta a través de un prisma.
- Entre 1670 y 1672, Sir Isaac Newton demostró que la luz blanca estaba formada por una
banda de colores: rojo, naranja, amarillo, verde, azul, añil/índigo y violeta. Lo descubrió
mediante la proyección de esta a través de un prisma.
- TEORÍA RETINEX
- En 1959, Edwin Land propone la teoría Retinex, partiendo de la constancia en la percepción del color a pesar de los cambios en la iluminación.
Esta teoría postula la existencia de tres sistemas independientes (sensibles a diferentes longitudes de onda) que forman imágenes que, aunque
nunca se mezclan, sí se comparan y se diferencian en claridad. De tal manera, cuando se combinan en la corteza cerebral, posibilitan la
percepción en color.
- En 1959, Edwin Land propone la teoría Retinex, partiendo de la constancia en la percepción del color a pesar de los cambios en la iluminación.
Esta teoría postula la existencia de tres sistemas independientes (sensibles a diferentes longitudes de onda) que forman imágenes que, aunque
nunca se mezclan, sí se comparan y se diferencian en claridad. De tal manera, cuando se combinan en la corteza cerebral, posibilitan la
percepción en color.
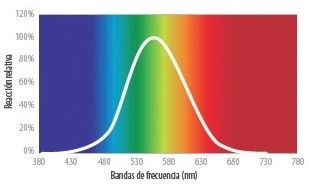
- LA CURVA DE SENSIBILIDAD FOTÓPICA
- Si la visión humana se expone a varias fuentes de luz monocromáticas de la misma intensidad o energía, la percepción del
brillo de esas fuentes no será el mismo, sino que dependerá de la longitud de onda. Este efecto se puede describir de esta
forma: si una determinada fuente de luz con un valor de energía y una cierta longitud de onda produce una determinada
sensación de brillo, para otra longitud de onda será necesario modificar esa energía para percibirla nuevamente con un brillo
igual a la primera. Esto da como resultado la curva de sensibilidad fotópica, y determina la sensibilidad relativa al brillo de la
visión humana según la longitud de onda.
- Esta curva nos muestra que el máximo de sensibilidad se produce aproximadamente en los
555 nm; otras longitudes de onda requieren más energía para producir la misma sensación de
brillo. La consecuencia natural de esta curva es que los colores extremos del espectro visible
(rojo y azul-violeta profundos) siempre se nos aparecerán como colores "oscuros" en
comparación con los verdes de igual energía.
- Esta curva nos muestra que el máximo de sensibilidad se produce aproximadamente en los
555 nm; otras longitudes de onda requieren más energía para producir la misma sensación de
brillo. La consecuencia natural de esta curva es que los colores extremos del espectro visible
(rojo y azul-violeta profundos) siempre se nos aparecerán como colores "oscuros" en
comparación con los verdes de igual energía.
- Si la visión humana se expone a varias fuentes de luz monocromáticas de la misma intensidad o energía, la percepción del
brillo de esas fuentes no será el mismo, sino que dependerá de la longitud de onda. Este efecto se puede describir de esta
forma: si una determinada fuente de luz con un valor de energía y una cierta longitud de onda produce una determinada
sensación de brillo, para otra longitud de onda será necesario modificar esa energía para percibirla nuevamente con un brillo
igual a la primera. Esto da como resultado la curva de sensibilidad fotópica, y determina la sensibilidad relativa al brillo de la
visión humana según la longitud de onda.
- EL OJO
Anexos de mídia
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Quer criar seus próprios Mapas Mentais gratuitos com a GoConqr? Saiba mais.