
49601
ADHD or ADD

Description
Module Information
Description
No tags specified
Show more
Show less
The incapacity of attending with a necessary degree of constancy to any one object (Sir Alexander Crichton, 1763–1856)
The first example of a disorder that appears to be similar to ADHD was given by Sir Alexander Crichton in 1798. Crichton was a Scottish physician who was born in Edinburgh in 1763. In 1785, he received his M.D. from the University of Leiden, The Netherlands (Palmer and Finger 2001; Tansey 1984). He then decided “to undertake a European medical tour” (Tansey 1984, p. 243) and practiced in hospitals in Paris, Stuttgart and Vienna (Tansey 1984). In his clinical practice, Crichton observed many cases of insanity and became increasingly interested in mental illness (Palmer and Finger 2001). In 1798, he published “An inquiry into the nature and origin of mental derangement: comprehending a concise system of the physiology and pathology of the human mind and a history of the passions and their effects”. In this work of three books, he demonstrated observations of clinical cases of mental illness (Palmer and Finger 2001). Up until the end of the eighteenth century, when Crichton published his inquiry, it was uncommon to focus on mental issues from a physiological or medical perspective (Palmer and Finger 2001). Crichton mentioned that at the time there were only two other authors who had “written fully on the subject of Mental Diseases” (Crichton 1798, pp. ii–iii, cited by Palmer and Finger 2001).
The second chapter of book II “On Attention and its Diseases” is of special interest to the present subject. Crichton begins this chapter with a definition of attention: “When any object of external sense, or of thought, occupies the mind in such a degree that a person does not receive a clear perception from any other one, he is said to attend to it” (Crichton 1798, reprint p. 200). Crichton emphasizes that the intensity of healthy attention varies within a normal range both between individuals and even within a person at different times (Crichton 1798). A distraction of attention does not necessarily have to be pathological, e.g. mental stimuli, volition, or education can have a great impact on healthy attention (Crichton 1798). Crichton distinguishes two possibilities of abnormal inattention as the oppositional poles of pathologically increased or decreased “sensibility of the nerves” (Crichton 1798):
The morbid alterations to which attention is subject, may all be reduced under the two following heads:
First. The incapacity of attending with a necessary degree of constancy to any one object.
Second. A total suspension of its effects on the brain.
The incapacity of attending with a necessary degree of constancy to any one object, almost always arises from an unnatural or morbid sensibility of the nerves, by which means this faculty is incessantly withdrawn from one impression to another. It may be either born with a person, or it may be the effect of accidental diseases.
When born with a person it becomes evident at a very early period of life, and has a very bad effect, inasmuch as it renders him incapable of attending with constancy to any one object of education. But it seldom is in so great a degree as totally to impede all instruction; and what is very fortunate, it is generally diminished with age. (Crichton, 1798, reprint p. 203)
Citation:(https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3000907/)
Description
No tags specified
Show more
Show less
Advances in the genetics of childhood neuropsychiatric disorders over the past decade were reviewed in the literature and reported from Yale University School of Medicine, New Haven, CT. In ADHD a genetic basis is suggested by family and twin studies. A focus on dopamine neurotransmission showed that children with ADHD had a higher incidence of the high-risk variant of DRD4 than controls, but the relationship is still controversial. The dopamine transporter gene (DAT1) has been linked to ADHD. Mice lacking expression of this gene were more active than control littermates. Functional variants of genes involved in dopamine transmission may confer a familial risk for ADHD. [1]
Parental ADHD. Weiss M, Hechtman L, and Weiss G provide a clinical perspective on ADHD in parents [2]. ADHD is highly familial. More than 50% of parents with ADHD have a child with ADHD, and 25% of children with ADHD have an ADHD parent. Siblings are frequently affected, and families with multiple ADHD members are especially challenged. Parents with ADHD are recognized for their failure to keep appointments for the child patient, they are restless in the office, they forget to bring along school reports, and they monopolize the interview. Parental ADHD can impact family functioning and the treatment of the child. Family counseling must address both the child’s and parent’s problems.
Executive functions and ADHD are reviewed by Barkley RA in Part 1 of a series of columns on the Genetics of ADHD [3]. The term executive function, deficient in ADHD, includes purposive, goal-directed activity; inhibition of distraction; response inhibition or delayed gratification; selective problem-solving; flexibility; goal persistence; and self-awareness. Self-regulation is essential for normal executive function. The prefrontal cortex is the anatomic localization of these functions. Attention deficits typical of ADHD are termed intention deficits.
Citation: ( Millichap, J.G., (2000). Genetics of ADHD. Pediatric Neurology Briefs. 14(8), pp.60–61. DOI: http://doi.org/10.15844/pedneurbriefs-14-8-7)
Heritability, family studies, suitability for genetic studies
Family and twin studies of cADHD demonstrate a high heritability, estimated to be around 70–80% from twin studies.6, 7 Relatively few studies have investigated the genetic and environmental contributions to the developmental course and outcomes in adulthood. Longitudinal twin studies show that the continuity of symptoms from childhood through to adolescence is predominantly due to common genetic influences.8, 9,10 Although such stable genetic effects are likely to continue beyond the adolescent years, there are only a few studies investigating this.
Genetic research on ADHD started with the finding that hyperactivity tends to aggregate in families.11, 12Since then, family studies have shown that ADHD shows familial clustering both within and across generations. Increased rates of ADHD among the parents and siblings of ADHD children have been observed.13, 14 In addition, strongly increased risks for ADHD (57%) among the offspring of adults with ADHD have been reported.15 Also, compared with the risk for ADHD among the siblings of children with ADHD (15%), siblings of adults with ADHD were found to have a strongly increased ADHD risk (41%).16Furthermore, a prospective 4-year follow-up study of male children into mid-adolescence found the prevalence of ADHD to be significantly higher among the parents and siblings of persistent ADHD child probands compared with the relatives of ADHD probands in whom ADHD remitted.17 Taken together, these studies suggest that the risk for ADHD may be greater among the first-degree relatives of probands with ADHD that persists into adolescence and adulthood than that among the relatives of probands with ADHD that remits before adulthood.17, 18
Whether such familial risks reflect genetic or environmental factors can be clarified using adoption and twin studies. Adoption studies found that ADHD is transmitted only to biological relatives, which strongly implicates genetic factors as the main causal influences on familial risk for the disorder.11, 12, 19, 20, 21These studies showed (for both current and retrospective symptoms in adults) that cADHD in child relatives predicts aADHD (or associated symptoms) in adult relatives. However, both adoption and family studies identify discrepancies related to different sources of ratings, with self-evaluation of ADHD symptoms by adults providing less evidence of familial effects than informants or cognitive performance data.19, 22, 23
Recently, four adult population twin studies using self-ratings of ADHD symptoms have been completed, which all found heritabilities that are far lower than those found in similar studies of parent- or teacher-rated cADHD: 41% for retrospectively reported childhood ADHD symptoms in a sample of 345 US veterans aged 41–58 years old,24 40% for current inattention problems in a Dutch study of 4245 18–30-year olds,8 30% for current ADHD symptoms in a Dutch study of over 12 000 twin pairs with an average age of 31 years25 and 35% for current ADHD in a Swedish sample of more than 15 000 twin pairs aged 20–46 years (Larsson et al., unpublished data). The situation is similar in adolescence, as adolescent twin studies using self-ratings show lower heritability estimates than studies of parent or teacher ratings,26, 27suggesting that self-ratings may be a poorer measure of the underlying genetic liability to ADHD than informant reports or clinical interviews. Although the estimated heritability in self-rated ADHD symptoms in adult populations is lower than that derived from parent or teacher ratings of cADHD, the pattern of findings is identical. Both types of studies find that there are no gender differences observed in the estimates of heritability, heritability estimates are stable across the age-span (for each type of measurement approach), there are similar estimates of the genetic correlation (the proportion of shared genetic effects) of 60–70% between inattention and hyperactivity-impulsivity, familial effects are all genetic in origin with no shared environmental influences, and no threshold effects are found. This suggests that for both child and adult ADHD the disorder is best perceived as the impairing extreme of a quantitative trait (Larsson et al., unpublished data; ref. 28).
Citation: (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3449233/)
Description
No tags specified
Show more
Show less
ADHD predominantly inattentive presentation
Fails to give close attention to details or makes careless mistakes
Has difficulty sustaining attention
Does not appear to listen
Struggles to follow through with instructions
Has difficulty with organization
Avoids or dislikes tasks requiring sustained mental effort
Loses things
Is easily distracted
Is forgetful in daily activities
ADHD predominantly hyperactive-impulsive presentation
Fidgets with hands or feet or squirms in chair
Has difficulty remaining seated
Runs about or climbs excessively in children; extreme restlessness in adults
Difficulty engaging in activities quietly
Acts as if driven by a motor; adults will often feel inside as if they are driven by a motor
Talks excessively
Blurts out answers before questions have been completed
Difficulty waiting or taking turns
Interrupts or intrudes upon others
Citation:(http://www.chadd.org/understanding-adhd/for-adults/diagnosis-of-adhd.aspx)
Description
No tags specified
Show more
Show less
Background
Pathophysiology of attention-deficit hyperactivity disorder (ADHD) is not known, and therefore the present study investigated mitochondrial defects, if any in cybrids created from patients and control population.
Methods
To investigate mitochondrial pathology in ADHD, cybrids cell lines were created from ADHD probands and controls by fusing their platelets with ρ0-cells prepared from SH-SY5Y neuroblastoma cell line. Cellular respiration, oxidative stress, mitochondrial membrane potential and morphology were evaluated employing oxygraph, mitochondria-specific fluorescence staining and evaluation by FACS, and immunocytochemistry. HPLC-electrochemical detection, quantitative RT-PCR and Blue Native PAGE were employed respectively for assays of serotonin, mitochondrial ATPase 6/8 subunits levels and complex V activity.
Results
Significantly low cellular and mitochondrial respiration, ATPase6/8 transcripts levels, mitochondrial complex V activity and loss of mitochondrial membrane potential and elevated oxidative stress were observed in ADHD cybrids. Expression of monoamine oxidizing mitochondrial enzymes, MAO-A and MAO-B levels remained unaffected. Two-fold increase in serotonin level was noted in differentiated cybrid-neurons.
Conclusions
Since cybrids are shown to replicate mitochondrial defects seen in post-mortem brains, these observed defects in ADHD cybrids strongly suggest mitochondrial pathology in this disorder.
General significance
Mitochondrial defects are detected in ADHD cybrids created from patients' platelets, implying bioenergetics crisis in the mitochondria could be a contributory factor for ADHD pathology and/or phenotypes.
Keywords: Cybrids, Mitochondrial membrane potential, ATPase, Respiration, Serotonin, Mitochondrial pathology
Go to:
1. Introduction
Inattention, impulsiveness and hyperactivity depict phenotypes of attention-deficit hyperactivity disorder (ADHD), a heritable and heterogeneous childhood disorder with a prevalence of 6-7% in children and adolescents [1]. Mitochondrial dysfunction is a suggested vulnerability factor in the pathogenesis of various neuropsychiatric disorders [2], [3], [4], [5]. Bioenergetic crisis during brain development, mitochondrial DNA (mtDNA) mutation or deletion may cause neurodevelopmental disorders. Autism spectrum disorder probands with mitochondrial defects exhibit symptoms of ADHD [6], [7], [8], suggesting possibility of bioenergetics defects in ADHD too [9]. Other than a report on significant stimulant effect of the ADHD drug, methylphenidate on neuronal firing and mitochondrial transport chain (ETC) enzyme activities in rats [10], no report is available on direct involvement of mitochondrial defects in ADHD. We investigated involvement, if any of mitochondrial bioenergetics in the pathophysiology of ADHD, by creating control and ADHD cybrids from an Indian population.
Cybrids, the mitochondrial transgenic cells created by fusion of mitochondria-less neuronal ρ0-cell lines with blood platelets from patients, mimic pathological conditions of disease phenotypes of post-mortem brains as shown for Parkinson’s disease [11] and other diseases [12]. We created cybrids using blood platelets from ADHD probands and ethnically-matched controls, and investigated their mitochondrial status and functions. One-time creation of cybrids has advantage over repeated blood sampling from young volunteers, since cybrids could be cryopreserved, differentiated into neurons and used continually for investigations.
Go to:
2. Materials and Methods
2.1. Materials
The human neuroblastoma cell line SH-SY5Y was obtained from National Centre for Cell Sciences, Pune, India. Low glucose Dulbecco’s modified Eagle’s medium, minimal essential medium modified for suspension culture (S-MEM), fetal bovine serum, Gentamicin, tetramethylrhodamine, methyl ester (TMRM), were procured from GIBCO, Invitrogen Corporation (CA, USA). Lymphocyte separation medium (LSM), polyethylene glycol 1000, manitol, were purchased from MP Biomedicals (France). PicoGreen®, MitoSOX™ were purchased from Molecular probes (OR, USA). Potassium chloride, KH2PO4, NaCl and Na2HPO4.2H2O, magnesium sulfate, lead nitrate, sucrose, were procured from Sisco Research laboratories (India). Amphotericin B, pyruvate, uridine, ethidium bromide, TRI reagent, 6-aminocaproic acid, Bis-Tris HCl, N-Dodecyl β-D-Maltoside, bis acrylamide, ATP, EGTA were purchased from Sigma (MO, USA). Coomassie Brilliant Blue G-250 and R-250 dyes were purchased from BioRad (CA, USA) and acrylamide was purchased from Merck (Darmstadt, Germany).
2.2. Methods
2.2.1. Cybrid preparation
ADHD probands (two aged 13; one aged 10 years) and ethnically-matched controls (a 5 years-old female, three males- 5 years, 6 years 8 months and 9 years old respectively) were recruited by mental health professionals of Manovikas Kendra following psychological evaluations through Conners’ Parents and Teachers Rating Scale [13] and Wechsler’s Intelligence Scale for Children [14] for assessing inattention/hyperactivity and Intelligence Quotient (IQ) levels, respectively. Patients with other neuropsychiatric disorders, pervasive developmental disorders, mental retardation (IQ > 70) including Fragile-X syndrome were excluded. Informed written consent for participation was obtained from the parents. Institutional Human ethics committee approved the study protocol.
2.2.2. Confirmation of mtDNA transfer
ρ0-Cells prepared from SH-SY5Y cell line were fused with platelets isolated from 3 ADHD male probands and two healthy controls to produce cybrids [15]. Genomic-DNA isolated from SH-SY5Y, ρ0-cells and cybrids created were subjected to long template PCR using internal primers that amplify a 5.8 kb fragment present in both nuclear and mitochondrial genomes and the amplicon was nested with external primers to amplify a 5.9 kb mtDNA-sequence [16]. PicoGreen® and Mitotracker Green® (Invitrogen Corporation, CA) staining were used for examining absence and presence of mtDNA in these cells and to locate mitochondria in live cells, respectively. Images of cells stained with PicoGreen® or Mitotracker Green® were captured using confocal microscope.
2.2.3. Mitochondrial membrane potential and oxidative stress
1 × 106 for fluorimetry and 2 × 105 for flowcytometry) of cells were plated in 6 well plates for TMRM staining. Next day cells were trypsinized and the pellet was resuspended in fresh medium containing 50 nM TMRM, incubated at 37 °C for 30 min in the dark in a CO2 incubator. The medium containing the stain was then removed and the cells were washed twice with D-PBS. Red fluorescence was measured using a spectrofluorimeter (PerkinElmer, USA). The excitation and emission wavelengths for TMRM staining were 530 nm and 570 nm respectively. Membrane potential was also measured with the same dye by using flowcytometry in cybrids. Cells were processed in the same way and the population of TMRM stained cells was analysed by flowcytometry (BD LSRFortessa (Mitochondrial membrane potential was evaluated by staining cells with tetramethylrhodamine, methyl ester (TMRM). Equal number fluorescence activated cell sorting (FACS), Software-FACS Diva 6.2.).
Cells were stained with MitoSOX™ to assess mitochondrial superoxide generation in these cybrids. Cybrids were collected by trypsinization and stained with 2.5 μM of MitoSOX™ for 30 min at 37 °C in the dark in a CO2 incubator. The stain containing medium was then removed and the cells were washed twice with D-PBS. Red fluorescence was measured using a spectrofluorimeter (PerkinElmer, USA, excitation: 510 nm, emission: 580 nm). MitoSOX™ stained cells were also analysed by flowcytometry (BD LSRFortessa fluorescence activated cell sorting (FACS), Software-FACS Diva 6.2.) [19].
2.2.4. Respiration
Whole cell respiration and digitonin permeabilized cells’ mitochondrial respiration were measured in cybrids using Oxygraph respirometer (Hansatech Instruments Ltd, England). Cells were trypsinized and added to the counter chamber of the Oxygraph respirometer containing D-PBS and the rate of oxygen consumption was measured for 8 min. For mitochondrial respiration the cells were permeabilized with digitonin (0.01%), in mitochondria isolation buffer (MIB, 225 mM Manitol, 75 mM Sucrose, 5 mM MOPs, 1 mM EGTA, dissolved in water, pH 7.4). The cells were washed two times to remove traces of digitonin and oxygen consumption was measured for 10 min. The representation of the amount of oxygen consumed was given as nmol of oxygen/min/number of cells use. [17].
2.2.5. Transcriptome analysis of ATP6/8 subunits
For real time PCR, RNA was isolated using TRI reagent according to the manufacturer’s protocol. Five μg of the total RNA was reverse transcribed using MuLV reverse transcriptase (conditions: 70 °C for 5 min, 37 °C for 5 min, then 42 °C for 60 min, and 70 °C 10 min, final hold at 4 °C). Relative quantification was performed using real-time PCR (Thermal Cycler Dice Real Time System TP800, Takara, Japan) with 100 ng of cDNA and SYBR Premix. The thermal cycling involved initial denaturation of 95 °C for 5 min followed by 40 cycles of 95 °C for 20 s, 58 °C for 30 s, 72 °C for 20 s. 18sRNA was used as endogenous control. The primers were designed using Primer 3 software (ATPase 6 - Forward 5'-GCCCTAGCCCACTTCTTACC-3', Reverse 5'-TTAAGGCGACAGCGATTTCT-3'; ATPase 8 - Forward 5'-CACCTACCTCCCTCACCAAA-3’, Reverse 5'-CTAGGATTGTGGGGGCAAT-3', 18srRNA – Forward 5’-CATGGCCGTTCTTAGTTGGT-3’, Reverse 5’ CGGACATCTAAGGGCATCAC-3’), data were analyzed using the 2-∆∆CT method [18].
2.2.6. Complex V activity
For Blue Native - Polyacrylamide gel electrophoresis (BN-PAGE), mitochondria (P2 fraction) were isolated in mitochondria isolation buffer (MIB, 225 mM Manitol, 75 mM Sucrose, 5 mM MOPs, 1mM EGTA, dissolved in water, pH 7.4), after quantification stored in -800C. Next day pellet was dissolved in sample buffer (1 M 6-aminocaproic acid and 50 mM Bis-Tris HCl, dissolved in water, pH 7.0), along with freshly prepared 10% (w/v) N-Dodecyl β-D-Maltoside, kept on ice for 10 min to dissolve the membrane proteins in the solution. Centrifuge the samples at 20,000xg for 30 min at 40C. Collect the supernatant and add the gel loading buffer (5% w/v Coomassie Brilliant Blue G-250 dissolved in 1 M 6-aminocaproic acid).
Gradient gel (5-13%) was prepared by using 30% acrylamide solution (by dissolving 29.22 g of acrylamide, 0.78 g of bis acrylamide in 100 ml of distilled water) in gel buffer (150 mM Bis-Tris, 1.5 mM aminocaproic acid, pH 7.0). The sample protein (60 μg) was run at, 100 V for 4 hrs at 4 0C on this gradient gel using 1X Cathode Buffer (10X Cathode Buffer- 50 mM Tricine, 15 mM bis-Tris HCl, pH 7.0, 0.02% Coomassie G-250) and 1X Anode buffer (10X Anode buffer- 500 mM Bis-Tris HCl, pH 7.0). Two gels were run using same protein, once run was complete one gel was kept for coomassie staining (for protein quantification, 0.25 % coomassie brilliant blue R250 in 40% methanol, 7% acetic acid) for one hour and distained overnight in 10% acetic acid and 10% methanol. Another gel was kept for complex V reaction buffer (35 mM tris, 270 mM glycine, 14 mM magnesium sulfate, 0.2 % lead nitrate, 8 mM ATP, pH 7.8) for overnight at room temperature. Next day gels were photographed, reaction gels shows white bands [19].
2.2.7. Serotonin analysis
For HPLC analysis 5 × 105 cells were plated and differentiated for six days. On the seventh day, the medium was replaced with fresh medium. After 16 h the cells were washed, sonicated in 50 μl of ice-cold 0.4 M perchloric acid, centrifuged at 12,500 rpm for 5 min and 10 μl of the supernatant was injected into the HPLC-ECD system. A standard solution containing 4 pmol of all the biogenic amines (Norepinephrine, DOPAC, Dopamine, 5-HIAA, HVA and 5-HT) were assayed prior to and at the end of the sample injections to confirm the retention time of the analytes in the column. The flow rate was 0.7 ml/min and the electrochemical detection was performed at 0.74 V. The sensitivity of the HPLC-ECD was set at 20 nA [21].
Cybrids cultured on poly-L-lysine coated cover-slips at 105 cells/ml/35 mm dish were immunostained for monoamine oxidase-A (MAO-A) and MAO-B proteins, images were captured employing confocal microscope, and fluorescence Intensity was measured using Image J software (NIH, USA) (Data are not shown)
Go to:
3. Results
3.1.1. Inheritance of mitochondria
A). In case of SH-SY5Y, control and ADHD cybrids PCR product of external and internal primers is obtained, while in ρ0 cells lacking mitochondria, only nuclear sequence amplified by internal primers was observed. PicoGreen dye binds to minor groove of Fig. 1The inheritance of mitochondria was tested in ADHD samples employing long-template PCR method. The primer sets were designed for PCR so that the internal primers amplified a 5.8 kb sequence present in both the nuclear (chromosome 1) and mitochondrial genomes and its amplicon was nested in that of the external primers, which amplified a 5.9 kb mtDNA-specific sequence (double stranded DNA and gives green fluorescence, so we can see the punctate mitochondria in the cytoplasm of SH-SY5Y cells. The punctate fluorescence is absent in ρ0 cells, but show only nuclear fluorescence because they do not contain mtDNA (Fig. 1B). The punctates observed in control, ADHD cybrids confirm their inheritance of mitochondria from platelets of healthy individuals and patients.
Fig. 1
Long-template PCR analysis and PicoGreen®staining of cybrids, ρ0-cells and SH-SY5Y cells: (A)Genomic DNA from SH-SY5Y, ρ0-cells, control and ADHD cybrids were amplified using both internal and external primers and subjected to electrophoresis on 0.6% agarose gel. Amplicons are absent in the PCR using external primers for ρ0 cells. (B) Punctated cytoplasm and nuclei are fluorescent in SH-SY5Y cells, control and ADHD cybrids, whereas in ρ0 cells only the nuclei exhibited green fluorescence by Pico-green staining. C6, C9, C10 and C11 are control cybrids, ADHD1, 5 and 6 are cybrids prepared using platelets of ADHD probands. Scale bar represents 10 μm.
3.1.2. ADHD cybrids show reduced mitochondrial mass and membrane potential
Mitotracker Green enters mitochondrial matrix where it covalently reacts with free thiol groups of cysteine residues of mitochondrial proteins and produce green fluorescence which allows visualization of the mitochondria. SH-SY5Y cells are showing elongated mitochondria and ρ0 cells have fragmented mitochondria (Fig. 2A). Bar diagrams represents the fluorescence intensity in the control and ADHD cybrids measured using Image J software which shows significant reduction in ADHD cybrids compared to controls (Fig. 2B). Mitochondrial membrane potential was examined in cybrid neurons with TMRM staining followed by spectrofluorimetric (Fig. 2C) quantification as well as flow cytometric (Fig. 2E and F) procedure. Significantly low TMRM fluorescence observed in ADHD cybrid-neurons implied higher mitochondrial depolarisation in these cells. Mitochondrial membrane potential was also analysed in differentiated cybrid neurons [20] by fluorimetric and FACS analysis and we observed similar results (data are not shown) as in undifferentiated neurons.
Open in a separate window
Fig. 2
Mitotracker Green®,TMRM and MitoSOX™ staining of control and ADHD cybrids: (A) Confocal images of Mitotracker Green® staining of the cybrids. (B) The intensity of Mitotracker Green® fluorescence quantified by Image J software. (C & D) The graphs showing the percent change of TMRM and MitoSOX™ fluorescence staining measured by fluorimetry and (E & G) FACS analysis, in ADHD as compared to the control cybrids. (F & H) are the representative scan of cybrids. Data are presented as Mean ± SEM. Values of p* ≤ 0.05 are considered significant. C6 and C9 are control cybrids, ADHD1, 5 and 6 are cybrids prepared using platelets of ADHD probands. Scale bar represents 10 μm.
3.1.3. Higher mitochondrial oxidative stress in ADHD cybrids
Likewise MitoSOX™ staining was carried out to test the mitochondrial oxidative stress generated in these cybrids neurons employing spectrofluorimetric (Fig. 2D) and flow cytometric (Fig. 2G and H) analyses. Interestingly ADHD cybrids displayed higher levels of superoxide radicals, as evidenced from the significantly increased MitoSOX™ fluorescence in the cells.
3.1.4. Reduced oxygen consumption due to complex V deficiency in diseased cybrids
Oxygen consumption was monitored in controls and ADHD non-permeabilized (Fig. 3A and B) and permeabilized cells (Fig. 3C and D) employing a sensitive Oxygraph in order to understand the respiratory capability of the cybrids. Significantly reduced rate of respiration found in ADHD cybrids, as well as in the mitochondria of these disease-cybrids, pointed to a significant loss of mitochondrial functions in both these diseases. Lower respiration rate could be resulting from the considerable loss in the ATPase 6/8 transcript levels in ADHD cybrids revealed in quantitative PCR analysis as seen in the present study (Fig. 3E and F).
Fig. 3
Whole cell and mitochondrial state3 respiration, APTase6/8 subunits expression, complex V activity and serotonin levels in control and ADHD cybrids: Whole cell respiration (A), cellular rate of oxygen consumption per 3 × 106 cells (B), mitochondrial State3 respiration (C), and mitochondrial oxygen consumption rate per 3 × 106 cells in ADHD and control cybrids (D). The cDNA prepared from total RNA of ADHD and control cybrids were subjected to qRT-PCR analysis using specific primers to accurately measure the transcript level of ATPase6/8 (E, F). Levels of serotonin in differentiated control and ADHD cybrid-neurons (G). Complex V activity using BN-PAGE, Activity gel (H), Coomassie staining gel (I). Bar diagram shows the difference in complex V activity analysed by Image software (J). (Control samples- 1 to 4 lanes and ADHD samples- 5 to 7 lanes in both the gels, M lane is marker). Data are presented as Mean ± SEM. Values of p* ≤ 0.05 is considered significant.
3.1.5. Higher level of 5HT in ADHD cybrid neurons and reduced activity of complex V
Serotonin level, measured by HPLC-ECD in differentiated neurons, was found to be higher in ADHD cybrids as compared to controls (Fig. 3G). BN-PAGE in-gel activity studies demonstrated a significant deficiency in mitochondrial complex V activity in ADHD cybrids, when compared to the control cybrids (Fig. 3H to J).
Go to:
4. Discussion
Most important feature of the present study is the successful creation of control and ADHD cybrids from an east-Indian population. This is confirmed by successful incorporation of mtDNA into ρ0-cells, which did not have mtDNA before fusion with the patient or control platelets. This is the first ever report on production of mitochondrial cell-hybrids (cybrids) for the neurodevelopmental disorder, ADHD. Another important attribute of this study is the finding of significant mitochondrial aberrations and dysfunctions, as evidenced by reduced staining intensity of Mitotracker Green®, TMRM and increase in MitoSOX fluorescence, and significant reduction in oxygen consumption concurrent with low levels of ATPase 6/8 transcripts in ADHD cybrids in comparison to control cybrid-neurons. These observations support a significant bioenergetic crisis in ADHD, which was proposed [9], but never demonstrated. The third important observation made in the present study is the significant increase in the levels of serotonin in the differentiated ADHD cybrid-neurons as compared to control cybrid-neurons, which is a topic of interest and intense debate.
Mitotracker green dye accumulates in the mitochondrial matrix where it covalently reacts with free thiol groups of cysteine residues and produce green fluorescence and allows to visualize the mostly inter-netted, tubular fluorescing mitochondria in the cytoplasm of control cybrids, but punctate (due to fragmented mitochondria) fluorescence in diseased cybrids. Quantification of mitotracker green staining by ImageJ software gives the total mass of the mitochondria present in the cytosol [23] which was found less in ADHD cybrids as compared to the control cybrids. This data speculates that due to fragmentation of mitochondria the total mass in the cytoplasm decreases. Significantly low TMRM fluorescence observed in ADHD cybrid-neurons implied higher mitochondrial depolarization in these cells. No report is available in literature which deals with mitochondrial membrane potential and ADHD, and therefore the present report is the first in literature. ADHD cybrids displayed higher levels of superoxide radicals, as evidenced by the significantly increased MitoSOX™ fluorescence in the cells. These amply provide proof that in ADHD there exists a serious oxidative stress condition, and that could be detrimental to the health of the neurons. In one study (48 children and adolescents (34 male, 14 female) with ADHD who had no neurological, systemic, or comorbid psychiatric disorders, and 24 sex- and age-matched healthy controls (17 male and seven female), it was observed that ADHD patients have oxidative imbalance [24]. In one another study, one group investigated the total antioxidative status and total oxidative status of plasma and antioxidant enzyme. Oxidative stress index values and the plasma TOS levels of the patients with ADHD were statistically higher than those of the control group [25]. Sezen et al. [26] also investigated and found that oxidative stress was higher in the ADHD patients than the control group. Bioenergetic dysregulation playing key roles in many neuropsychiatric disorders is recently reviewed [27]. This may result from defective mitochondrial ETC complex function, which is validated by a significant decrease in state3 respiration in ADHD cybrids. A considerable decrease in the ETC complex V ATPase 6/8 subunits in ADHD cybrids further provides positive evidence in this direction. ATPase 6/8 mutations are associated with many neurodevelopmental disorders including autism [28], [29], with which ADHD shows overlapping syndromes [30]. The decreased transcript level of ATP6 and ATP8 subunits may lead to the lower activity of complex V in ADHD cybrids as our BN-PAGE data shows.This evidence firmly suggest that mitochondrial dysfunction exists in ADHD.
The status of mitochondrial energetics in ADHD subjects is not available. However, indirect evidences from pharmacological studies using methylphenidate, which is a drug commonly used to treat ADHD have shown that the drug affects the activity of mitochondrial ETC enzymes and results in increase of the extracellular levels of the catecholamines that stimulate glycolysis and release of lactate from the astrocytes, thereby correcting the energy deficiency, and restoring appropriate firing rates [10]. This has led to the hypothesis that impairment of mitochondrial bioenergetic machinery may play a role in autism and ADHD pathogenesis [9], [31].
Serotoninergic system, known to modulate impulse control and aggression, is challenged in ADHD patients [32]. At a time when mitochondrially-located MAO-A or MAO-B were unaffected in ADHD cybrids, the differentiated cybrid-neurons from ADHD probands exhibited a 2-fold increase in serotonin indicating probably an active role of this biogenic amine in the phenotypes of the disease. If this is proved to be true, these cybrid-neurons can be used as a cellular model for ADHD to investigate neuromolecular mechanisms underlying ADHD pathology.
At a time when no direct evidence of mitochondrial dysfunctions in ADHD is available, we created and used ADHD cybrids to assess mitochondrial functions in this neurodevelopmental disorder. Cybrids created by the fusion of ρ0-cells and Parkinson’s disease (PD) patient platelets are shown to replicate mitochondrial pathology seen in post-mortem PD brains [11], and differentiation of these cells into neurons are demonstrated to retain the mitochondrial defects [22]. Therefore, ADHD cybrids and the cybrid-neurons created in the present report could be reliable tools to study the mitochondrial dynamics and bioenergetics and to understand the molecular mechanisms underlying the pathophysiology of ADHD.
In short, this is the first report that identifies mitochondrial dysfunction as an inherent factor of ADHD pathology, and for a probable direct role of the serotoninergic system in the disease syndromes. Increased serotonin content in the differentiated cybrid-neurons suggests that the mitochondrial defect in these cells can significantly alter the serotonergic neurotransmitter function, which could be a pathological hallmark of ADHD. This study identifies mitochondrial pathology as one of the risk factors for ADHD, and the hybrid model provides a novel, reliable and valid tool to study the molecular basis of ADHD at a cellular level.
Citation: (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5121149/)
Description
No tags specified
Show more
Attention Deficit Disorder is one of the most well-recognized childhood developmental problems.
But it doesn’t end in childhood. Did you know that a full 60% of those diagnosed with ADD in childhood will continue to have issues that affect their functioning as adults? It’s true. And ADD can fly under the radar in adults who were never diagnosed in childhood.
While prevalence rates are not entirely clear for adults, as mentioned earlier, they are estimated to fall within the 4% to 5% range.
That’s a lot of adults struggling with attention problems! What’s more, ADD remains one of the most misunderstood and incorrectly treated illnesses today.
But that is changing.
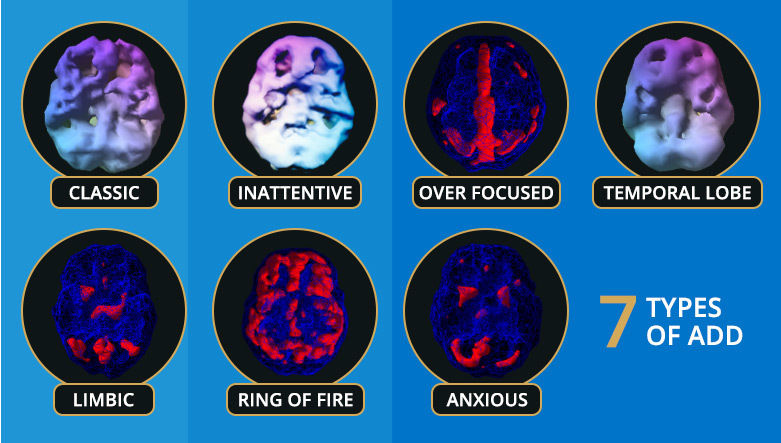
Using breakthrough diagnostic techniques, Dr. Amen has discovered that there are 7 distinct types of ADD, each requiring a different treatment.
This is good news because more people can be helped with treatments that are tailored to their specific ADD type.
But the first step in healing from ADD is to determine if you have it.
Here are several identifying behaviors to look for – and the 7 types of ADD revealed:
There are five longstanding hallmark symptoms of ADD:
Short attention span. People with ADD have a difficult time with boring tasks (such as regular, routine, everyday tasks) and need stimulation or excitement in order to stay engaged. Many people with ADD can pay attention just fine for things that are new, novel, interesting, highly stimulating, or frightening.
Distractibility. People with ADD tend to notice more in their environment than others, which makes them easily distracted by outside stimuli, such as light, sounds smells, certain tastes or even the clothes they wear. Their keen sensitivity causes them to get easily off task.
Disorganization. Most people with ADD tend to struggle with organization of time and space. They tend to be late and have trouble completing tasks on time. Many things get done at the last moment or even later. They also tend to struggle keeping their spaces tidy, especially their rooms, book bags, filing cabinets, drawers, closets and paperwork.
Procrastination. Tasks and duties get put off until the last moment. Things tend not to get done until there are deadlines or someone else is mad at them for not doing it.
Poor internal supervision. Many people with ADD have issues with judgment and impulse control and struggle not to say or do things without fully thinking it through. They also have a harder time learning from their mistakes.
You need at least three of these symptoms over a long period of time, and these symptoms, in some way, should interfere with your life.
Here’s a brief description of each type:
Type 1. Classic ADD (ADHD) – inattentive, distractible, disorganized, hyperactive, restless, and impulsive.
Type 2. Inattentive ADD – inattentive, easily distracted, disorganized, and often described as space cadets, daydreamers, and couch potatoes. Not hyperactive!
Type 3. Overfocused ADD – inattentive, trouble shifting attention, frequently get stuck in loops of negative thoughts or behaviors, obsessive, excessive worrying, inflexible, frequent oppositional and argumentative behavior. May or may not be hyperactive.
Type 4. Temporal Lobe ADD – inattentive, easily distracted, disorganized, irritable, short fuse, dark thoughts, mood instability, and may struggle with learning disabilities. May or may not be hyperactive.
Type 5. Limbic ADD – inattentive, easily distracted, disorganized, chronic low grade sadness or negativity, “glass half empty syndrome,” low energy, tends to be more isolated socially, and frequent feelings of hopelessness and worthlessness. May or may not be hyperactive.
Type 6. Ring of Fire ADD – inattentive, easily distracted, irritable, overly sensitive, cyclic moodiness, and oppositional. May or may not be hyperactive.
Type 7. Anxious ADD – inattentive, easily distracted, disorganized, anxious, tense, nervous, predicts the worst, gets anxious with timed tests, social anxiety, and often has physical stress symptoms, such as headaches, and gastrointestinal symptoms. May or may not be hyperactive.
Citation:(https://www.brainmdhealth.com/blog/how-to-identify-attention-deficit-disorder-in-yourself-or-a-loved-one-discover-the-7-types-of-add/)
Show less
Media
{kind=link}
Context
Description
No tags specified
Show more
Show less
Treating ADHD often requires medical, educational, behavioral and psychological intervention. This comprehensive approach to treatment is sometimes called “multimodal” and, depending on the age of the individual with ADHD, may include:
parent training
medication
skills training
counseling
behavioral therapy
educational supports
education regarding ADHD
Working closely with health care providers and other professionals, treatment should be tailored to the unique needs of each individual and family to help the patient control symptoms, cope with the disorder, improve overall psychological well-being and manage social relationships.
Citation:(http://www.chadd.org/understanding-adhd/about-adhd/treatment-of-adhd.aspx)