1575880
Beschreibung
Mindmap von Kristi Brogden, aktualisiert more than 1 year ago
|
|
Erstellt von Kristi Brogden
vor etwa 10 Jahre
|
|
Maps without frontiers
- Mapping differences
- Visual system
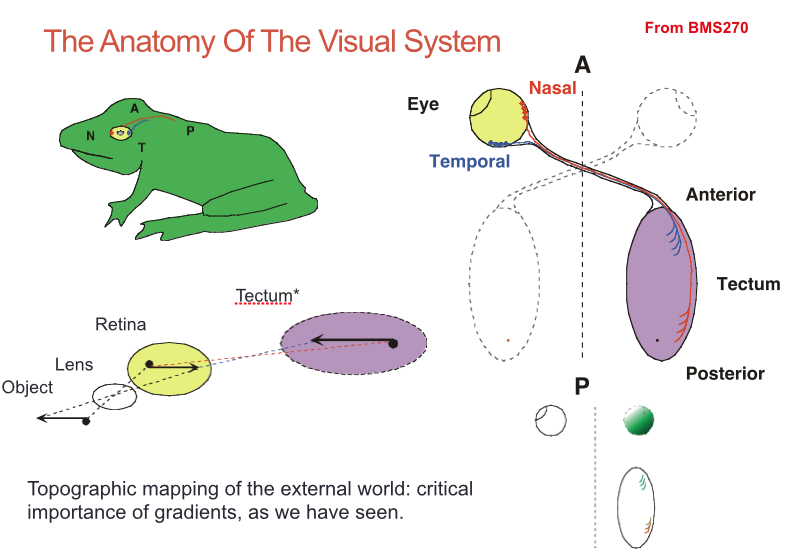
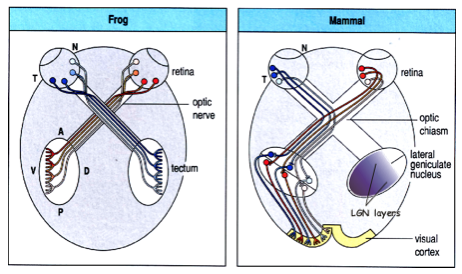
- In addition to projections to the tectum (superior
colliculus), mammalian retinal ganglion cells (RGCs)
also project to the lateral geniculate nucleus (LGN),
which relays these inputs to the cortex:
- Interestingly, the mapping of RGC axons onto the LGN is
also topographic and is set up using similar gradients of
Ephrins
- (Feldheim et al., 1998,
Neuron, 21 1303)
- Unlike the tectum, the LGN
receives inputs from both eyes
- allows stereoscopic
vision to be integrated
- Terminals from, say, the temporal retina of the left eye
(blue) are located in adjacent layers of the same region of
the LGN as those from the temporal right eye (red)
- Guidance of RGC axons to
these layers is predetermined.
- Guidance of RGC axons to
these layers is predetermined.
- allows stereoscopic
vision to be integrated
- (Feldheim et al., 1998,
Neuron, 21 1303)
- Interestingly, the mapping of RGC axons onto the LGN is
also topographic and is set up using similar gradients of
Ephrins
- In addition to projections to the tectum (superior
colliculus), mammalian retinal ganglion cells (RGCs)
also project to the lateral geniculate nucleus (LGN),
which relays these inputs to the cortex:
- Olfactory system
- How do you represent a
non-spatial sense?
- 1000 receptors but each
neuron expresses only one!
- “One neuron –
one OR” principle
- “One neuron –
one OR” principle
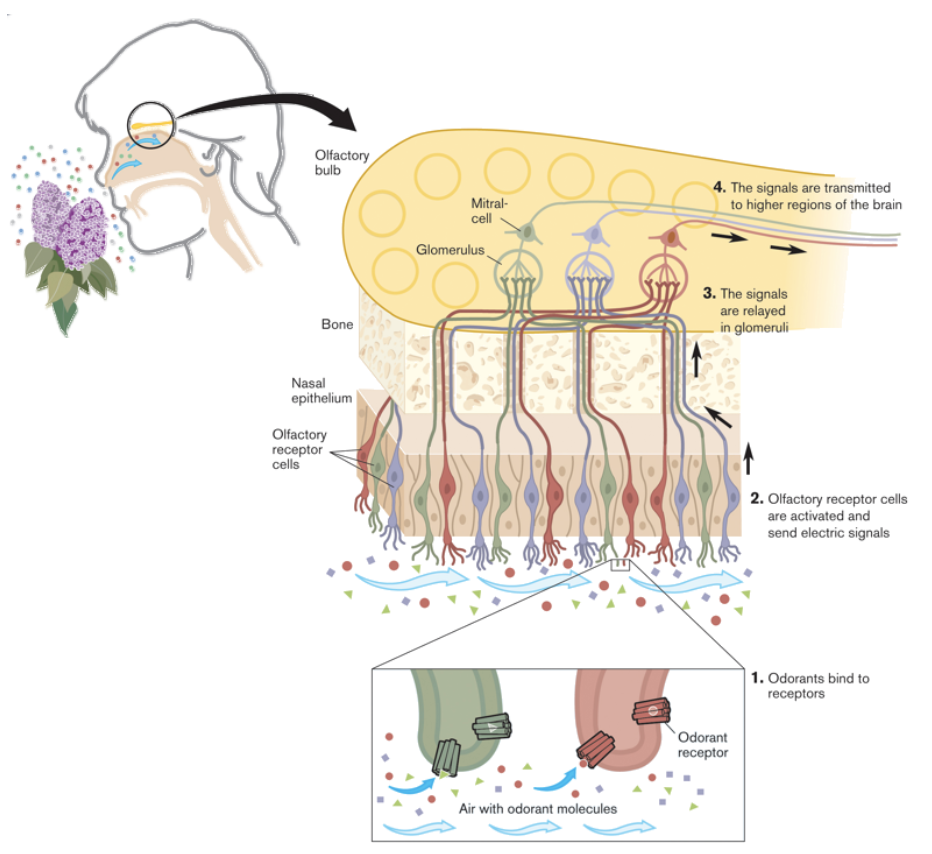
- Receptor expression dispersed in nasal epithelium, but
axons become organised in the olfactory bulb (OB)
- How is this done?
- Mapping from
epithelium to bulb
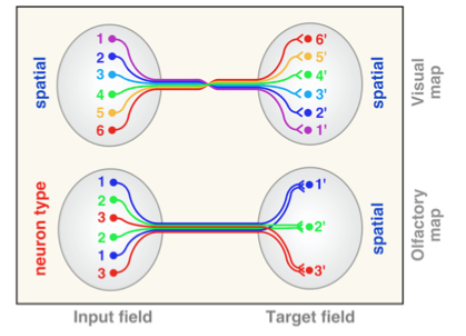
- Critically different
from RT mapping:
- Glomeruli responding
to related odorants
are clustered in OB
- Critically different
from RT mapping:
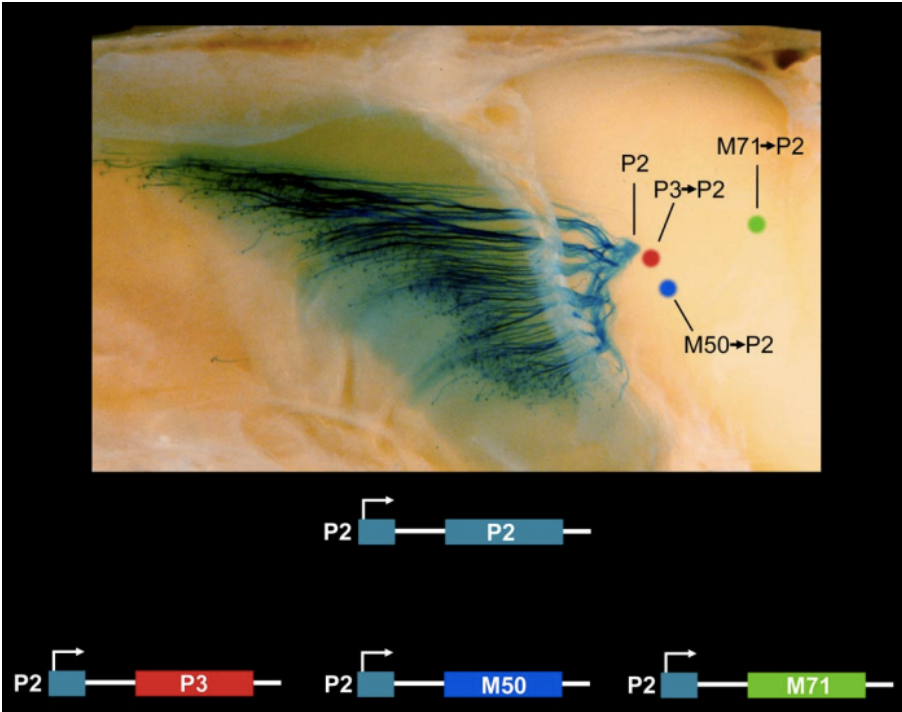
- Receptor expression governs guidance
- Receptor swap experiments demonstrate
that where axons go is determined by
which receptor is expressed.
- OR activity determines
guidance response
state
- Olfactory receptors (ORs) are 7 TM
GPCR-like molecules
- In the absence of ligand (odour), each
receptor has a characteristic basal activity.
Early guidance is activity-independent.
- In the absence of ligand (odour), each
receptor has a characteristic basal activity.
Early guidance is activity-independent.
- Neurons expressing the same OR
have similar cAMP signalling levels
(adenylate cyclase dependent;ACIII)
- This determines the level of transcription
of familiar guidance cues (Robo/Slit,
Eph/Ephrin, Neuropilin/Sema).
- This results in receptor/cue protein
levels characteristically associated
with expression of a particular OR,
which, in turn, determines mapping
in olfactory bulb (OB)
- Disruption of guidance cue
expression (e.g. Neuropilin2)
disrupts regional mapping in OB
- This results in receptor/cue protein
levels characteristically associated
with expression of a particular OR,
which, in turn, determines mapping
in olfactory bulb (OB)
- This determines the level of transcription
of familiar guidance cues (Robo/Slit,
Eph/Ephrin, Neuropilin/Sema).
- Olfactory receptors (ORs) are 7 TM
GPCR-like molecules
- Receptor swap experiments demonstrate
that where axons go is determined by
which receptor is expressed.
- Conversion from
continuous to discrete map
- Axons entering OB are
pre-sorted due to
cue/receptor interactions
- Cue expression switches with time (e.g. from
Robo to Nrp/Sema) so that early entering axons
then guide later entering axons.
- Cue expression switches with time (e.g. from
Robo to Nrp/Sema) so that early entering axons
then guide later entering axons.
- Sorting into glomeruli is
activity-dependent
- Activity drives higher cAMP levels which turns on
expression of homophilic adhesion molecules (Kirrels
and contactins), and Ephs and Ephrins (again!)
- These interactions sort axons
expressing same ORs into groups
to form the glomeruli.
- These interactions sort axons
expressing same ORs into groups
to form the glomeruli.
- Activity drives higher cAMP levels which turns on
expression of homophilic adhesion molecules (Kirrels
and contactins), and Ephs and Ephrins (again!)
- Axons entering OB are
pre-sorted due to
cue/receptor interactions
- Mapping from
epithelium to bulb
- And, does this mean there is
also spatial organisation of
olfactory info in the cortex?
- How is this done?
- 1000 receptors but each
neuron expresses only one!
- How do you represent a
non-spatial sense?
- Visual system
- Olfactory
system
experiments
- Choi et al. Cell (2011) vol. 146 (6) pp. 1004-15
- Experimental strategy
- Introduce ‘channelrhodospin’ (ChR2)
into subset of PC neurons
- ChR2 is a light-activated cation
channel that stimulates action
potentials upon exposure to light
- ie can ‘fire’ PC neurons
independent of mitral
cell input.
- ie can ‘fire’ PC neurons
independent of mitral
cell input.
- Stimulate the ChR2+ subset of
neurons with light, paired with
either an aversive or appetitive
(appetite inducing) stimulus in
naïve (unconditioned) animals
(classic associative learning).
- After conditioning, test whether light
stimulus alone can elicit the
appropriate behavioural response.
- After conditioning, test whether light
stimulus alone can elicit the
appropriate behavioural response.
- Introducing
channelrhodopsin
to PC neurons
- 3 ways (all using viruses)
- Simplest: use
synapsin
promoter
- Hits 50% of cells
at injection site
- Hits 50% of cells
at injection site
- Infect floxed* Chr2 into mouse in
which cre driven from Emx1
promoter (excitatory
neuron-restricted)
- * These lox sites are in ‘flip’
orientation so cre will invert
the gene not delete it
- Also hits 50%, but
only excitatory
neurons
- * These lox sites are in ‘flip’
orientation so cre will invert
the gene not delete it
- Infect floxed* Chr2 at same
time as virus containing
synapsin driving cre
- Much lower Chr2
expression rate
(10%)
- Much lower Chr2
expression rate
(10%)
- Simplest: use
synapsin
promoter
- 3 ways (all using viruses)
- ChR2 is a light-activated cation
channel that stimulates action
potentials upon exposure to light
- Introduce ‘channelrhodospin’ (ChR2)
into subset of PC neurons
- ChR2 activation
can condition
aversive behaviour
- Photostimulation (PS) of ChR2-expressing neurons
in the piriform cortex - the conditioned stimulus
(CS) - was paired with foot shock – the
unconditioned stimulus (US) - on only one side of
the chamber to condition the animals (10 pairings).
- Animals then exhibited flight behaviour to
PS alone, but only when ChR2 was present
in piriform neurons (and a minimum of 200
had to be infected with ChR2).
- Conditioning with odorants and PS together,
showed that subsequently either PS or
odorants could elicit flight.
- Animals then exhibited flight behaviour to
PS alone, but only when ChR2 was present
in piriform neurons (and a minimum of 200
had to be infected with ChR2).
- Photostimulation (PS) of ChR2-expressing neurons
in the piriform cortex - the conditioned stimulus
(CS) - was paired with foot shock – the
unconditioned stimulus (US) - on only one side of
the chamber to condition the animals (10 pairings).
- ChR2 photostimulation can also
drive appetitive behaviours
- Mice trained to take water in
response to odorant, could be
re-trained to respond instead
to PS
- Male mice also could be trained to
associate presence of a female with either
an odour as the CS or with PS as the CS
- Mice trained to take water in
response to odorant, could be
re-trained to respond instead
to PS
- Piriform cortex neurons are
plastic in their associative
capability
- The same set of
ChR2-expressing PC neurons
can be re-trained in either
direction
- Distinct sets of
ChR2-expressing PC neurons
can be trained and retrained
to elicit different behaviours
- ie the piriform cortex is a
very plastic substrate
- The same set of
ChR2-expressing PC neurons
can be re-trained in either
direction
- Does this prove that the
PC is the site of odorant
learning?
- No, just shows that PC
can be used for
associative learning.
(What would prove it?)
- However, does show that PC is very
plastic: apparently any group of ~200
neurons can be used to elicit diverse
behavioural associations, reversibly.
- NB similar experiments in other regions of
the cortex (e.g. somatosensory) elicited a
specific behavioural output according to
location (ie topographically constrained).
Huber et al., 2008 Nature v451, p61
- NB similar experiments in other regions of
the cortex (e.g. somatosensory) elicited a
specific behavioural output according to
location (ie topographically constrained).
Huber et al., 2008 Nature v451, p61
- Nonetheless, strongly
suggests that random
connections from OB into
PC are used to associate
odours with particular
experiences.
- No, just shows that PC
can be used for
associative learning.
(What would prove it?)
- Experimental strategy
- Choi et al. Cell (2011) vol. 146 (6) pp. 1004-15
- Responses to
odorants
- Learned
- Piriform cortex
- Beyond the bulb…
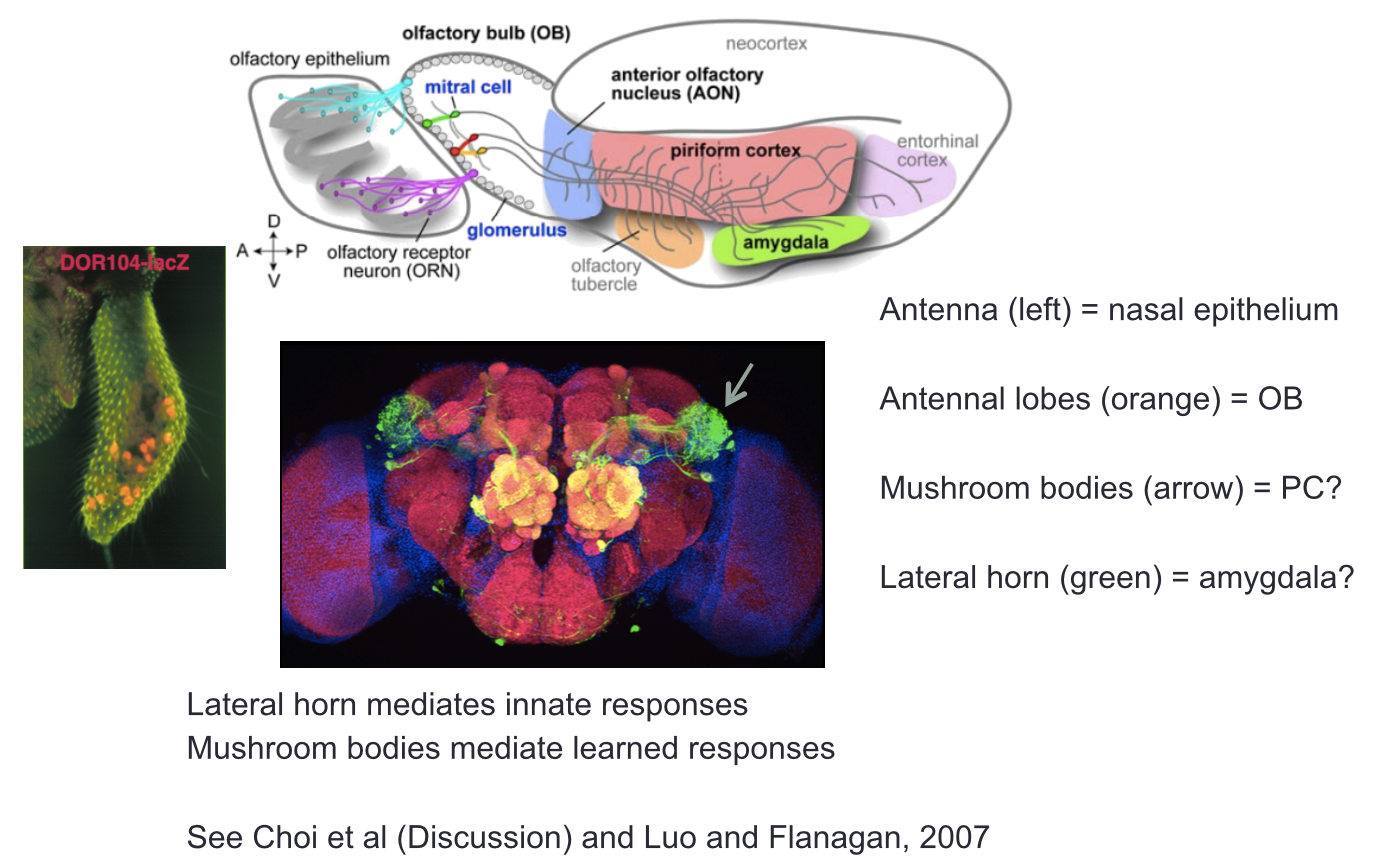
- As in the visual system, olfactory signals are relayed
from the bulb to multiple higher centres (e.g. SC,
LGN, then to cortex in visual system)
- However, unlike the visual system, mitral cell
axons projecting to the piriform cortex (PC) do
not exhibit any spatial organisation
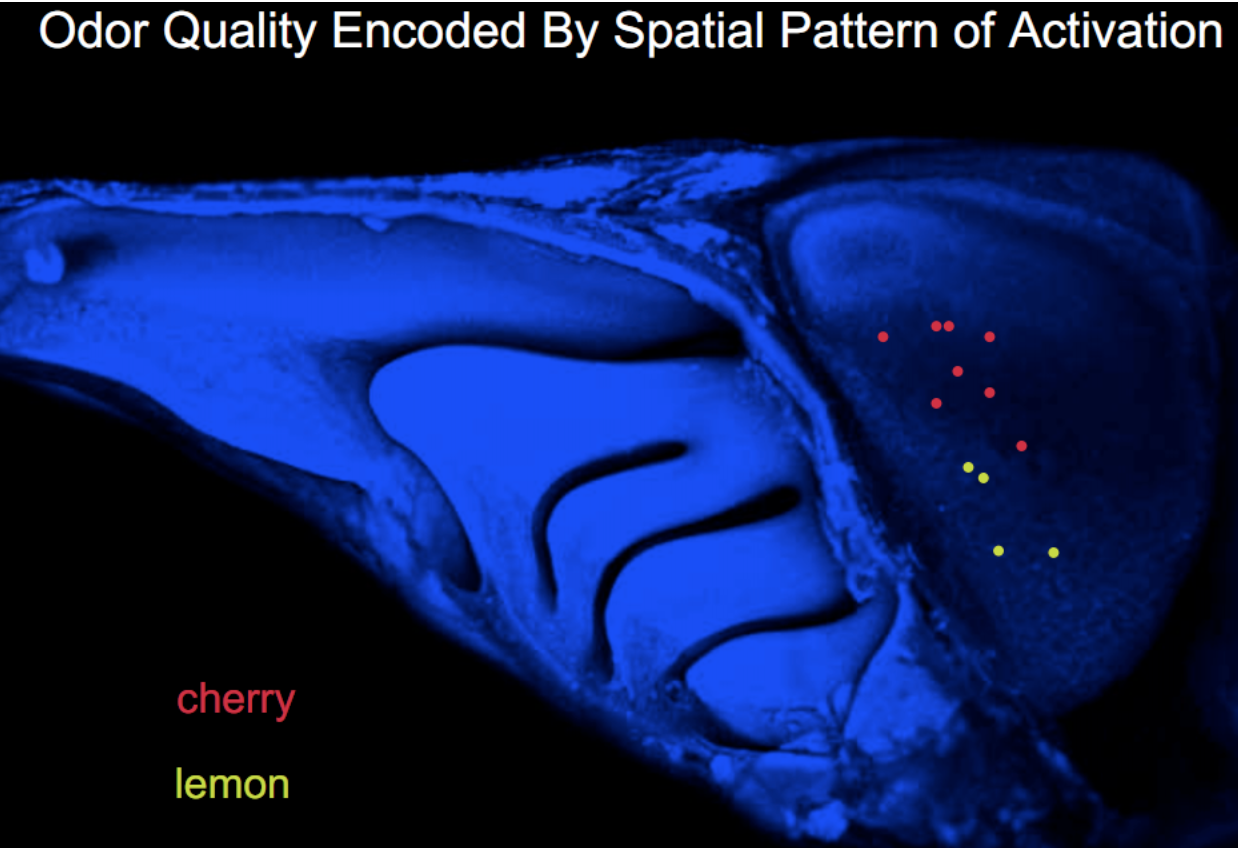
- Correspondingly, individual odorants activate
subpopulations of neurons distributed across
the PC
- NB Individual PC neurons respond to multiple, structurally dissimilar odorants
- NB Individual PC neurons respond to multiple, structurally dissimilar odorants
- Correspondingly, individual odorants activate
subpopulations of neurons distributed across
the PC
- However, unlike the visual system, mitral cell
axons projecting to the piriform cortex (PC) do
not exhibit any spatial organisation
- As in the visual system, olfactory signals are relayed
from the bulb to multiple higher centres (e.g. SC,
LGN, then to cortex in visual system)
- Is the piriform cortex the
site of olfactory learning?
- Choi et al., (2011) test this using
optogenetic activation of arbitrary
subsets of PC neurons…..
- Choi et al., (2011) test this using
optogenetic activation of arbitrary
subsets of PC neurons…..
- Beyond the bulb…
- How does the brain
know which
odorant is which?
- In mammals, the majority of
odors only drive behaviour after
learning. ie the significance of
odors is learnt by association.
- However, it is not
known which brain
regions are involved.
- However, it is not
known which brain
regions are involved.
- Piriform cortex
- Innate
- Are all responses to odorants learned?
- A small subset of odours elicit innate responses
- e.g. trimethyl-thiazoline (TMT) from fox elicits fear (in mice!)
- There are spatially invariant
projections from OB to cortical
amygdala that may be involved.
- Similar bifurcation in flies……
- There are spatially invariant
projections from OB to cortical
amygdala that may be involved.
- e.g. trimethyl-thiazoline (TMT) from fox elicits fear (in mice!)
- A small subset of odours elicit innate responses
- Drosophila olfactory
system: conserved function
- Are all responses to odorants learned?
- Learned
Medienanhänge
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Möchten Sie kostenlos Ihre eigenen Mindmaps mit GoConqr erstellen? Mehr erfahren.