Beschreibung
|
|
Erstellt von Richard Duport
vor mehr als 9 Jahre
|
|
Seite 1
Gynécologie - 2 - Stéroïdes et cycle menstruel
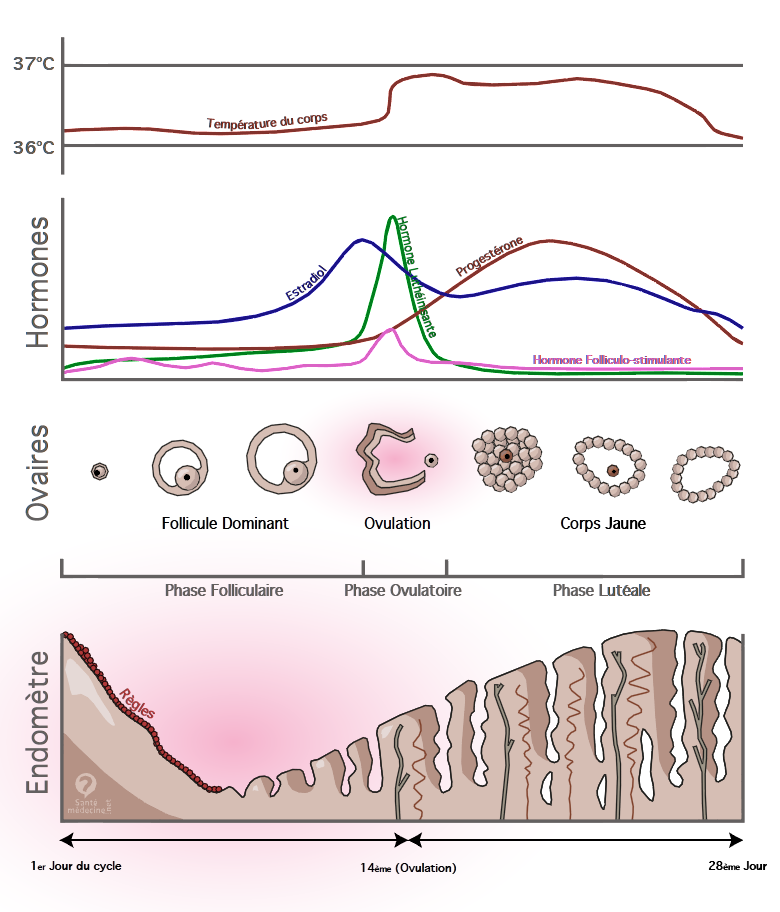
1. Les hormones stéroïdiennes sexuelles Stimiulation: par des glycoprotéines FSH et LH, de taille élevée --> récepteur membranaire --> AMPc (2nd messager)Transport: Testostérone et Oestrogènes: 69% liés à SHBG (foie); 30% à albumine Progestérone: 80% liée à l'albumine et 18% liée à la CBG (et non SHBG comme indiqué dans les dias!) Fracitons libres: 1% d'esotrogènes et de testostérone, 2% de progestérone --> effet tampon et préservation de la dégradation 1 site unique de liaison sur SHBG pour oesotrgènes ou androgènes Taux d'insuline et BMI inverses aux taux d'eoestrogènes liés, au profit des oestrogènes libres --> lien entre surpoids et pathologies endométriales Effets sur la cellule-cible: diffusion passive: --> cytoplasme --> noyau --> récepteur --> trasncription d'un ARNm --> protéine (messager final)Excrétion: métabolisation dans le plasma: oestrone ou oestriol (oestrogènes faibles) --> conjugaison --> désactivation stéroïdienne et hydrosolubilité--> urines et un peu voies biliairesBiosynthèseC27: Cholestérol --> C21: dérivés pregnane et corticostéroïdes --> C19: dérivés androstane puis androgènes --> C18 --> dérivés estranes ou oestrogènes.Cholestérol apportés principalement à partir des LDL (50%) (rappel= chylomicrons, seulement 10%; VLDL surtout TG; HDL surtout phospholipides et prot., protège CV)LDL: apolipoprotéines B avec récepteur spécialisé à la surface des ovaires. Si saturé --> récepteurs périphériques au niveau de l'intima artérielle --> athérosclérose.Pré et post-ménopause: HDL augmenté de 10mg/dL --> fréquence des problèmes CV plus élevée chez l'homme. LDL augmentent après ménopause: 1ère cause de décès chez la femme de plus de 50 ans = problèmes CV Enzymes: Communes dans l'ovaire et la surrénale, sauf 21-hydroxylase (corticostéroïdes) Surrénales: surtout voie "delta-4" des hormones sexuelles (à partir de la progestérone): androsténedione; Ovaires: aussi "delta-5": DHEA. Le système du cytochrome P450 P450SCC (mitochondries des cellules thécales) = desmolase --> clive chaîne latérale de 6 atomes de carbone --> dérivés pregnane P450C17 (réticulum endoplasmique) --> androgènes (thèque mais pas granulosa!) P450 aromatisant (réticulum endoplasmique) --> oestrogènes (activité cent fois supérieure dans la granulosa que dans la thèque) 2. Le double-système cellulaire Rappels histologiques: Les follicules passent pas les stades primordial, pré-antral, antral et préovulatoire. Au stade primordial, quelques cellules stromales forment la thèque, autour du follicule primordial. La granulosa apparaît au stade pré-antral comme un épaississement cellulaire autour d'une membrane pellucide qui entoure le follicule. Les stades antral et préovulatoire est caractérisé par une accumulation de liquide pellucide (riche en stéroïdes). Contact intime constant entre thèque et granulosa. Première partie du cycle ovulatoire. LH se fixe à son récepteur membranaire sur les cellules de la thèque --> AMPc --> stimulation du cytochrome P450 --> synthèse d'androstènedione et de testostérone. Ces androgènes diffusent passivement vers la granulosa. FSH se fixe à son récepteur membranaire sur les cellules de la granulosa --> AMPc --> stimulation du cytochrome P450arom --> synthèse d'oestrone et d'oestradiol Les oestrogènes diffusent passivement --> thèque --> plasma Deuxième partie du cycle post-ovulatoire Persistance du système bi-cellulaire évoqué ci-dessus. Persistance de la synthèse d'androgènes et d'oestrogènes. Mais en plus, FSH stimule la synthèse de récepteurs à LH à la surface des cellules de la granulosa --> synthèse de progestérone. Rq: la granulosa n'a pas de système P450C17 et dépend donc de la thèque pour la synthèse d'androgènes. Facteurs régulateurs IGF II stimule synthèse d'androgènes par la thèque et d'oestrogènes par la granulosa Inhibine inhibe la synthèse de FSH mais stimule l'activité de P450C17 et donc la synthèse d'androgènes. 3. Les hormones Métabolisme Les oestrogènes sont synthétisés par les ovaires (oestrone E1et oestradiol E2) et les tissus périphériques adipeux, musculaires et cutanés (oestrone). Les surrénales et les ovaires sont les sources d'androstènedione. 1% seulement sera converti en périphérie. 100 à 300 µg/j d'oestradiol 3µg d'androstènedione sont synthétisés par jour par l'ovaire. La conversion par les tissus périphériques: tissu adipeux, muscles, peau forme 45µG/J d'oestrone La progestérone est fabriquée à raison d'1mg/j en pré-ovulatoire et de 20-30 mg/J en phase lutéale. Elle est dégradée en pregnanediol, alors que la 17-OH-progestérone est dégradée en pregnanetriol, tous deux éliminés dans les urines. La DHEA provient à 50% des surrénales, 25% de l'ovaire et 25% des tissus périphériques. L'androstènedione provient à 50% des surrénales et 50% des ovaires. La testostérone provient à 25% des surrénales, 25% des ovaires et 50% de conversion périphérique --> 0,2 - 0,3 mg/j Son métabolite actif est la DHT qui nécessite la 5-alpha-réductase (intracellulaire). Mécanisme d'action Récepteur intracellulaire: cytoplasmique (androgènes) ou nucléaire (oestrogènes) La diffusion se fait passivement, donc selon le gradient de concentration Toutes exercent un rétro-contrôle négatif sur l'hypothalamus (GnRH) Oestrogènes L'oestradiol a une plus forte affinité que l'oestrone pour son récepteur, mais l'action oestrogénique dépend aussi de la quantité d'homorne présente et la durée de l'exposition (ex: personne en surpoids fabrique beaucoup d'estrone pendant longtemps --> effets importants) Deux types de récepteurs aux oestrogènes: ER-alpha et ER-beta, de prévalences variables selon le tissu Ex: endomètre: effets alpha prolifératifs vs beta anti-prolifératifs. Il existe des co-récepteurs et des co-activateurs selon les tissus agissant sur les facteurs de transcription TAF-1 ou TAF-2. Tissus ayant des récepteurs aux oestrogènes: organes génitaux, SNC (psycho-dynamique, augmente activité électrique --> épilepsie), glande mammaire, adipocytes, os, granulosa ovarienne, peau, foie, système CV, 75% des cancers mammaires. Les effets sont principalement anabolisants; cas particuliers: rétro-contrôle sur l'hypothalamus (GnRH), adipocytes abdominaux inhibés au profit des glutéofémoraux; augmente la coagulation et la fibrinolyse. Structure du récepteur aux oestrogènes (pas vu en cours) Domaine AB: action TAF 1 d'activation de la transcription; oestrogène-indépendante. Domaine C: région DNA-ligant --> reconnaît le gène cible Domaine D: région charnière (H pour Hinge): 1/ signal permettant passage vers noyau où se trouve l'hormone; 2/ modification de conformation du récepteur une fois lié à l'hormone. Domaine EF: région hormone-ligant; 1/ Fixation de heat shock-proteins pour éviter fixation à l'ADN tant que non fixé à l'hormone; 2/ TAF 2: activation de la transcription, oestrogène-dépendante. Exemple du Tamoxifène: inhibiteur compétitif des récepteurs oestrogéniques (SERM de 2ème génération) TAF-1 est indépendante du ligant et agoniste, mais TAF-2 est dépendante --> inhibée compétitivement. Antagoniste sur le sein mais agoniste sur l'os (protecteur) et l'endomètre (augmente de 1/2000 à 1/1000 le risque de cancer de l'endomètre; atrophie endométriale kystique et polypes endométriaux fréquents) Illustre la répartition des effets hormonaux selon les tissus et les co-inhibiteurs/activateurs présents. Progestérone Deux types de récepteurs à la progestérone: PRA et PRB Effets sur le SNC (hypothalamus), sein organes génitaux, métabolisme glucidique. Légèrement anabolisant, parfois synergique avec oestrogènes (morpho-différenciation) S'oppose aux oestrogènes sur les organes génitaux (diminution filance et abondance de la glaire cervicale, antiproliférative mais sécrétoire sur endomètre, relaxant sur le myomètre. Elève la T° (courbe ménothermique) Effets négatifs: augmente le risque de cancer mammaire en post-ménopause; SNC: hypnotique et dépressif; diminue la tolérance au glucose Androgènes Stimule agressivité, anabolisme protéique, obésité androïde, pilosité, raucité de la voix, glandes sébacées; Favorisent l'insulino-résistance et la chute du HDL-cholestérol 4. Cycle menstruel Les cycles qui suivent la ménarche et précèdent la ménopause sont anovulatoire et ont une durée beaucoup plus variable.Axe HHG Noyau arqué de l'hypothalamus postérieur: cellules neuronales peptidiques (neurones pouvant synthétiser des hormones peptidiques) GnRH relarguée dans le système porte hypothalamo-hypophysaire. 10 acides aminés, durée de vie très courte: 2-4 minutes. --> Stimule les cellules gonadotropes à FSH et LH de l'hypophyse antérieure sur trois niveaux: synthèse et mise en réserve; activation et transfert vers site de relargage; excrétion plasmatique. 3 boucles de feed-back: long (LH et FSH plasmatiques sur GnRH); court (LH et FSH sur GnRH via système porte); ultra-court (GnRH sur elle-même). Noradrénaline stimule le noyau arqué, la dopamine l'inhibe. Ces dernières sont modulées par les endorphines. Une inhibine est aussi synthétisée au niveau hypophysaire: effet autocrine --> inhibe synthèse et sécrétion FSH; stimule excrétion LH. Les 4 rétro-contrôles exercés par les hormones stéroïdiennes: 1/ [oestrogènes] faibles --> stimule synthèse et stockage FSH/LH mais inhibition de la sécrétion de FSH; peu d'influence sur sécrétion de LH. 2/ [oestrogènes] hautes ( > seuil critique ) --> stimule sécrétion ++ LH (cf. pic de LH) 3/ [progestérone] faibles --> stimule synthèse FSH (cf. pic de FSH après pic LH) 4/ [progestérone] hautes --> inhibent pulsatilité GnRH --> inhibition sécrétion gonadotrophines. Rq: la pilule est donc gonadostatique par rétro-contrôle négatif sur l'hypophyseLa maturation des follicules Migration cellules germinales primitives vers les crêtes germinales --> follicules primordiaux: mitoses jusque 16-20 semaines de gestation --> 6-7 millions Atrésie --> décroissance à 2 millions à la naissance, 300 000 à la puberté; 400 donneront lieu à une ovulation. Croissance sur 60-70 jours des follicules primordiaux puis accélération pendant 14 jours (phase pré-ovulatoire): 3 à 10 sont recrutés pour aller vers le stade pré-antral. La FSH est primordiale dans la préservation des follicules de l'atrésie. A partir du stade pré-antral: expression des récepteurs à la FSH au niveau de la granulosa qui grandit; FSH et oestrogènes stimulent l'expression de récepteurs à FSH --> auto-entretien et augmentation de la sensibilité aux oestrogènes. fragilité: si trop d'androgènes --> saturation des aromatases et le follicule devient androgénique et s'atrésie. Stade antral: activité bi-cellulaire à fond; mais toujours fragile: si trop de LH --> trop d'androgènes --> Atrésie. Sélection: résultat du combat entre l'auto-entretien qui renforce le follicule (oestrogènes et FSH stimulent la synthèse d'oestrogènes) et le rétro-contrôle négatif des oestrogènes sur la sécrétion de FSH. Le follicule dominant a un taux de récepteurs FSH élevé qui lui permet de survivre malgré la chute progressive de FSH. Inversion: le haut taux d'oestrogènes au niveau du follicule dominant modifie l'action de la FSH qui stimule la synthèse de récepteurs à LH dans les cellules de la granulosa (préparation à la fonction lutéale) Les hormones pendant le cycle menstruel (cf. image ci-dessous-Phase pré-ovulatoire: Noter d'abord que la température augmente pendant la deuxième partie du cycle sous l'effet des progestérones Au début du cycle, le follicule dominant est sélectionné autour de J5-J7. Le taux d'oestrogènes augmente progressivement exerçant un rétro-contrôle négatif sur la FSH. Cela contribue à l'atrésie des follicules non-dominants. En fin de phase folliculaire, la LH augmente progressivement, sous l'effet des oestrogènes en augmentation. 3 choses en pré-ovulatoire: stimulation par FSH de la synthèse de récépeteurs à LH au niveau granulosa et début de synthèse de progestérone avant oulation; rétro-contrôle négatif de la FSH sur elle-même via la synthèse d'inhibine par elle-même; l'inhibine stimule cependant l'androgénèse de la thèque Pic de LH: stimulus nécessaire à l'ovulation par inhibition de l'OMI (ovary maturation inhibitor) réenclenchement de la mitose Progestérone et LH favorisent la synthèse d'enzymes protéolytiques --> rupture du follicule. élimination définitive des follicules non-dominant; induction de la production de progestérone et entretien de la synthèse ses récepteurs; la progestérone inhibe les mitoses de la granulosa; favorise la production de prostaglandines le follicule. La thèque est stimulée par le pic de LH ainsi qu'atrésie des follicules non-dominants --> augmentation d'androgènes locaux et périphériques Pic de FSH La progestérone restaure le feedback positif des oestrogènes sur la FSH et permet donc le pic de FSH. Il permet la présence de récepteurs suffisants à la LH pour une phase lutéale normale. Il permet de libérer l'ovocyte de ses attaches au niveau du cumulus de la granulosa Il favorise la dégradation du plasminogène en plasmine puis en collagénases, favorisant le phénomène ovulatoire. Phase post-ovulatoire La lutéine donne la couleur jaune. Le corps jaune est très vascularisé ce qui permet l'apport en cholestérol. La durée de ce corps jaune est génétiquement pré-programmée à 14 jours. La granulosa produit de l'inhinbine --> maintien de FSH à taux bas. La progestérone agit aux niveaux local et central pour inhiber toute nouvelle croissance folliculaire. Si grossesse, par contre, HCG prend le relai jusqu'à stéroïdogénèse placentaire à 9-10 semaines. Régression du corps jaune La régression du corps jaune est mal expliquée: oesrogènes locaux seraient lutéolytiques? Conséquences: effondrement d'oestrogènesn, de progestérone (reprise pulsatilité GnRH) et d'inhibine (levée le rétro-contrôle négatif sur FSH) FSH reprend: recrutement de nouveaux follicules. 5.En pratique Pour évaluer la réserve ovulaire: on dose à J3 du cycle: FSH 45 pour une réserve correcte.Pour évaluer le caractère anovulatoire d'un cycle: on dose la progestérone à J22.A l'approche de la ménopause, on fait un test à la Progestérone en mettant 10 jours la patiente sous progestérone. A l'arrêt, la patiente devrait saigner, sauf si les oestrogènes sont trop bas pour la croissance de l'endomètre. Ca signe la ménopause.Test à la LHRH: explore la capacité sécrétoire de l'axe HHG. On injecte de la LHRH IV en phase folliculaire; la LH doit x3-5 à 30 minutes; la FSH x1,5-2,5 à 60 minutes.Test à la dexaméthasone: on administre 1mg de DXM à 23h la veille --> dosage de la cortisolémie et des hormones surrénaliennes qui doivent être supprimées. (Rappel: une faible dose (1mg) inhibe une sécrétion non pathologique; une forte dose (8mg) inhibe une sécrétion d'ACTH pituitaire, mais pas une sécrétion par un adénome surrénalien.
{kind=link}
Réponses aux objectifs du chapitre Reproduire les étapes biochimiques de synthèse des stéroïdes sexuels en identifiant les principales fonctions enzymatiques.Les stéroïdes sexuels sont synthétisés à partir du cholestérol, une molécule lipidique de 27 atomes de carbone. Les enzymes responsables des étapes de la synthèse des stéroïdes sexuels sont regroupées sous l'entité de cytochrome P450. La première réaction est un clivage de la chaîne latérale à 6 carbones par une desmolase, le P450SCC, une enzyme mitochondriale. Le résultat donne naissance aux molécules à 21 atomes de carbone: les dérivés pregnanes (progestatifs et corticostéroïdes). Ceci se produit dans le reticulum endoplasmique.C'est ensuite le complexe du P450C17 qui produit une nouvelle classe de molécules, l'androstane puis les androgènes, à 19 atomes de carbone.Enfin, une aromatisation par le P450arom permet de transformer ces dernières en molécules à 18 atomes de carbones, les estranes ou oestrogènes. Cela se produit de nouveau dans le reticulum endoplasmique. (paragraphe suivant: hors-sujet?)Il est à noter que cette synthèse suit des voies différentes et des portions propres en fonction de la cellule. Les cellules de la thèque interne sont synthétisent jusqu'aux androgènes sans pouvoir aromatiser ces derniers en oestrogènes. Ceci se fait des les cellules de la granulosa ovariennes, ou encore dan les tissus périphériques (peau, muscles, graisses). Les cellules de la granulosa sont quant à elles dépendantes de la thèque interne en androgènes puisqu'elles n'ont pas de P450C17. Les surrénales, pour leur part, produisent aussi des androgènes selon une voie "delta-4", qui aboutit à la formation de DHEA-S (spécifique), de DHEA et d'androstènedione. Elles peuvent aussi produire les minéralo- et gluco-corticoïdes grâce à la 21-hydroxylase, ces dernières n'étant pas des hormones sexuelles. Décrire le système bicellulaire de synthèse des stéroïdes et son aspect dynamique au cours du cycle menstruel.Le système bicellulaire ovarien est constitué des cellules des deux couches entourant le follicule: la thèque et la granulosa.Le substrat de cette synthèse, le cholestérol, arrive par voie sanguine porté par les LDL. Les cellules de la thèque disposent de l'attirail enzymatique nécessaire à la production d'androgènes, androstènedione et testostérone, à partir du cholestérol (P450SCC et P450C17). Ces derniers diffusent alors passivement, notamment vers les cellules de la granulosa qui possèdent des enzymes P450arom, absentes des cellules de la thèque, permettant d'aromatiser les androgènes en oestrogènes (oestrone et oestradiol). Il y a donc dépendance en substrats androgènes des cellules de la granulosa vis-à-vis des cellules de la thèque, pour fabriquer des oestrogènes.Au début du stade folliculaire, le follicule est primordial. Seule la thèque enveloppe le follicule et la synthèse de stéroïdes se limite aux androgènes, sous l'effet stimulant de la LH basale. L'élévation progressive de la FSH permet au follicule primordial d'échapper à l'atrésie et de se développer au stade pré-antral, une fine couche de granulosa l'entoure alors. Les androgènes formés dans la thèque diffusent passivement vers la granulosa où ils sont aromatisés en oestrogènes. La FSH et les oestrogènes stimulent la synthèse de récepteurs à a FSH, auto-entretenant ainsi la synthèse d'oestrogènesAu stade antral, le follicule est entouré de liquide folliculaire, riche en oestrogènes. Le système bicellulaire reste fragile, puisque tout excès d'androgènes mène à l'atrésie du follicule. Ceci peut être dû à une saturation du système d'aromatisation des cellules de la granulosa par exemple, et un taux de récepteurs à la FSH suffisant est nécessaire. C'est ce qui caractérisera le follicule dominant. Lorsque la concentration en oestrogènes atteint un seuil dans le follicule dominant, la FSH provoque une augmentation de la synthèse des récepteurs à la LH au niveau des cellules de la granulosa, démarrant la fonction supplémentaire, lutéale, de synthèse de progestérone dans ces cellules. Puis, le taux élevé d'oestrogènes circulant induit le pic de LH, qui déclenche l'ovulation (inhibition de l'OMI et stimulation de la synthèse et de la libération d'enzymes protéolytiques) et élimine définitivement les follicules non dominants via la poussée androgénique qui y fait suite dans la thèque. La progestérone à un taux faible permet le pic de FSH. Ce dernier contribue à libérer l'ovocyte de ses attaches et assure le maintien d'un nombre suffisant de récepteurs à la LH pour une phase lutéale normale.La phase lutéale est caractérisée par une vascularisation importante du corps jaune, pour un apport important de cholestérol associé à une synthèse conséquente de stéroïdes sexuels: androgènes, œstrogènes et progestérone. Le système bicellulaire est toujours actif, les cellules de la granulosa restant dépendantes en androgènes des cellules de la thèque pour la synthèse d'oestrogènes. La FSH est maintenue à un niveau bas par la synthèse de grandes quantités d'inhibine par les cellules de la granulosa.La perte de fonction du corps jaune se traduit par un effondrement du taux d'oestradiol, de progestérone et d'inhibine, ce qui lève le rétro-contrôle négatif sur la FSH et sur la pulsatilité de la sécrétion de GnRH. Ceci permettra un nouveau recrutement pour le cycle suivant.Décrire de manière détaillée le cycle menstruel et le cycle anovulatoireFaire un dessin du graphe du cycle avec FSH, LH, oestrogènes et progestérone.Au début du cycle menstruel, la progestérone, les oestrogènes et l'inhibine sont à des taux très bas. Le rétro-contrôle sur la FSH est levé et sont taux augmente progressivement. Ceci contribue au développement de quelques follicules (une dizaine typiquement) primordiaux. S'y développe alors la couche de cellule de la granulosa. Les androgènes synthétisés au niveau de la thèque sont alors aromatisés par les cellules de la granulosa en oestrogènes. La FSH et les oestrogènes stimulent la synthèse de récepteurs à la FSH, auto-entretenant ainsi le développement du follicule et sa sensibilité à la FSH.Les taux bas d'oestrogènes stimulent la synthèse et le stockage de FSH et de LH au niveau central, mais inhibe la sécrétion de FSH, dont le taux reste faible.Un follicule particulier se démarquera et développera une forte quantité d’œstrogènes. Les oestrogènes circulants vont permettre la croissance de l'endomètre.En fin de phase folliculaire, la FSH favorise la synthèse de récepteurs à la LH au niveau des cellules de la granulosa. Ceci permet la sécrétion par celles-ci de progestérone, dont le taux augmente déjà avant l'ovulation. Puis, le haut d’œstrogènes déclenche le pic de LH. Ce dernier permet l'ovulation par inhibition de l'OMI (ovulation maturation inhibitor) et stimulation de la synthèse de protéases. La concentration faible de progestérone induit la restauration d'un feed-back positif des oestrogènes sur la sécrétion de FSH, ce qui permet le pic de FSH. Ce dernier favorisera le maintien d'un nombre suffisant de récepteurs à la LH pour une phase lutéale normale. Le pic de LH aura aussi induit une hausse du taux local d'androgènes synthétisés par la thèque, qui confirmera l'atrésie des follicules non-dominants.Pendant la phase lutéale, le corps jaune est richement vascularisé et ainsi bien alimenté en substrat des hormones stéroïdiennes sexuelles, le cholestérol. La pulsatilité de la sécrétion de GnRH est abolie par le haut taux de progestérone. Ceci, ainsi que les taux élevés d'inhibine sécrétée dans la granulosa induit une inhibition de la synthèse, du stockage et de la sécrétion de gonadotrophines LH et FSH. La progestérone permettra la maturation de l'endomètre et sa préparation à l'implantation.En fin de phase lutéale, l'effondrement des oestrogènes, de la progestérone et de l'inhibine lèvent le rétro-contrôle sur la FSH. La sécrétion de GnRH retrouve sont caractère pulsatile. Ceci permet le recrutement de nouveaux follicules primordiaux et le début d'un nouveau cycle. La chute du taux de progestérone provoque l'atrophie de l'endomètre, bien vascularisé, entraînant les menstruations.En cas de cycle anovulatoire, le taux d'oestrogènes n'est jamais suffisamment élevé pour que le pic de LH ait lieu. La progestérone reste donc constamment à des taux bas. Il n'y a pas non plus de pic de FSH. Pour détecter le caractère anovulatoire d'un cycle, la solution idéale est de doser la progestérone au jour 22 du cycle, ou plutôt 6 jours avant les règles suivantes si le cycle ne dure pas régulièrement 28 jours.Enoncer trois test dynamiques permettant d'explorer l'axe hypothalamo-hypophyso-gonadique / surrénalien, et en expliquer les principes.Test à la LHRH: permet d'explorer la capacité de sécrétion en hypohysaire en gonadotrophines. On injecte 100µg de LHRH en IV pendant la phase folliculaire. On mesure la LH à 30 minutes (elle doit être multipliée par 3-5) et la FSH à 60 minutes (elle doit être multipliée par 1.5 à 3.5 par rapport à la valeur basale). Si ce n'est pas le cas, nous avons une insuffisance secrétoire.Test à la dexaméthasone: on donne 1mg de DXM per os le soir à 23h (car le pic d'ACTH se fait la nuit). On dose le lendemain matin le taux de cortisol et d'androgènes surrénaliens. Ceci teste l'axe hypothalamo-hypophyso surrénalien. Il doit y a voir inhibition de cet axe par rétro-contrôle négatif de la DXM et on doit donc mesurer des taux bas d'androgènes surrénaliens et de cortisol. Ce test est notamment utilisé pour diagnostiquer un syndrome de Cushing.Test aux progestatifs: il consiste à donner de la progestérone per os pendant 10 jours, pendant trois mois consécutifs. L'absence de saignements vaginaux au bout des 10 jours signe une hypooestrogénémie et donc la ménopause. Ce test se fait autour de l'âge de la ménopause, vers les 50 ans, par exemple en présence de bouffées de chaleur, quand on veut s'assurer que la ménopause est bien présente sans attendre un an d'aménorrhée.Test à J3 de la FSH: le but est d'évaluer la réserve ovulatoire. On dose au 3è jour du cycle la FSH plasmatique, à un moment du cycle où le taux d'oestrogènes circulants doivent être bas, induisant un rétro-contrôle négatif sur la FSH (il faut donc aussi doser l'oestradiol plasmatique). Pour qu'une réserve ovulatoire soit jugée correcte, on souhaite avoir une FSH inhibée à Enoncer les sources synthétiques des stéroïdes sexuels et leur contribution respective aux productions d'androgènes et d'oestrogènes.Ici le sens de la question est pas évident. Je donne une proposition de réponse sans grande certitude.Le cholestérol est le substrat primordial de toute hormone stéroïdienne sexuelle. Il est transformé en androgènes au niveau des surrénales et des cellules de la thèque ovariennes. Une partie de la testostérone est obtenue, toutefois, par conversion périphérique d'androgènes précurseurs. Leur aromatisation en oestrogènes peut se faire dans les cellules de la granulosa ovarienne ou des tissus adipeux.Les androgènes circulants sont l'androstènedione (50% surrénalienne, 50% ovarienne), la DHEA (50% surrénalienne, 50% ovarienne), la DHEA-sulfate (100% surrénalienne) et la testostérone (25% surrénalienne, 25% ovarienne, 50% périphérique, i.e. peau, muscles, tissus gras). Les oestrogènes circulants sont l'oestradiol (ovarien) et l'oestrone, qui est soit de source ovarienne, soit le produit de transformation d'oestradiol ou d'aromatisation d'androgènes circulants par les tissus gras.
Möchten Sie kostenlos Ihre eigenen Notizen mit GoConqr erstellen? Mehr erfahren.