16396627
Descripción

Fichas por Alice Hathaway, actualizado hace más de 1 año
|
|
Creado por Alice Hathaway
hace más de 5 años
|
|
| Pregunta | Respuesta |
| Why enzyme used? | Cell reactions not spontaneous or quick enough Enzyme don't change equilibrium position Allows rapid rate at biological conditions |
| Ribozymes | Made of RNA Rest of enzymes made from proteins |
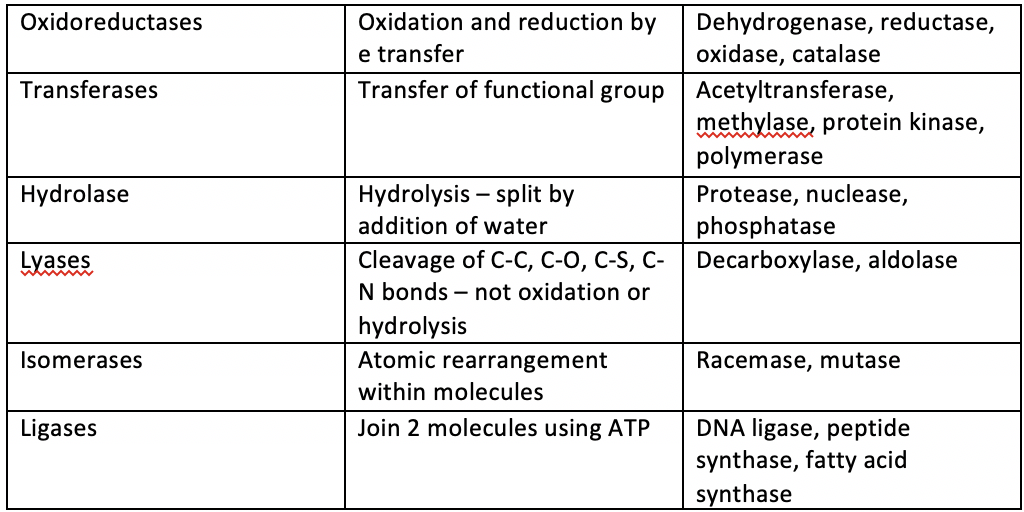
| Types of Enzyme | |
| Prosthetic group | Small molecule integral to structure Tightly bound Involved in catalysis |
| Cofactors | Small molecules, essential for enzyme activity Can be tightly bound prosthetic groups, or loosely bound cosubstrates - dissociated to be regenerated Can be ions or organic coenzymes |
| Coenzymes | Based on essential vitamins needed in diets E.g. electron carriers as amino acid side chains have no electron carrying capacity |
| How enzymes work | Active site binds to substrate Stereospecific Reversibel formation of ESC Can breakdown or form product Decrease activation |
| Transition state | Top of energy barrier Relatively unstable Energy change between substrate and transition state much lower in enzyme catalysis |
| Catalysis | Speed up rate at which equilibrium reached Alternative reaction pathway Acid base catalysis; covalent catalysis; proximity effects; substrate strain |
| Lock and key | IF true, ESC would be favourable hence product wouldn't form |
| Induced Fit | Subtle arrangement increase complementarity, placing strain hence enzyme binds more tightly to transition state than substrate or product |
| Rate depends on,,, | [Enzyme] [Substrate] |
| Km | Substrate concentration at half Vmax Higher - slower rate Michaelis Menten constant Intrinsc property of enzyme on substrate Includes how well enzyme catalysed formation of product after ESC formed |
| Michaelis menten equation | Vmax[s]/Km[s] |
| Kd | Dissociation constant (rate constant) If K3 small compared to K2, Km~Kd |
| K3 | Formation of product from ESC Also Kcat - turnover number |
| Kcat/Km | Measure of affinity Large ratio = efficient enzyme Rate limited by diffusion of substrate |
| Notable Kcat | Carbonic Ahydrase = 600,000/sec Lysozyme = 0.5/sec |
| Linwever Burk Equation | Reciprocal of Michaelis-menten gradient = Km/Vmax Y intercept = 1/Vmax X intercept = 1/Km |
| Units of activity | Katal 1 Katal = 1 molecule of substrate/ sec |
| How do enzymes work? | ESC increase local effective concentration fo reactants. Unfavourable loss of entropy when substrate binds offset by enthalpy change of binding |
| Features of enzymes | Denaturation destroys activity Stereospecific for substrate and cofactors Active site in hydrophobic clefts Enzyme interact with non-covalent interactions Proximity adn orientation of reactive groups of substrate important - stabilise transition state |
| Lysozyme | Natural antibiotic in sweat, egg white, tears, saliva Cuts peptidoglycan at peptide links, causing lysis |
| Structure of peptidoglycan links | NAG and NAM NAM linked with amino acid via peptide bond. D isomer hence not hydrolysed by protease |
| How lysozyme works | Binds to sugar moiety Hydrophobic active site accommodates 6 residues, all fit easily except D - strained Offset by favourable bidding of others, but forced into half chair hence more reactive Bond between D and E cut by enzyme |
| Amino acid action | Glu-35 and Asp-52 either side of bond between D and E Dissociate to form negatively charged carboxylate with neutral pH Glu-35 behave as acid (abnormal high pKA) Asp-52 deprotonated |
| Old mechanism | Glu-35 donates H to glucosidic bond, leaving ring D with positive charged carbon Stabilised by -ve charge of Asp-52 and lone pair O electrons. Carbon reacts with water activated by -ve change on Glu-35. Water donates OH to D and H to regenerate Glu-35 (Incorrect - instead nucleophilic attack) |
| New mechanism | Nucloephilic attack by Asp-52 on C1 of ring D displaces oxygen of glycosidic bond. Takes carbonyl hydrogen from glucose-35 glycosyl enzyme intermediate reacts with water to form second product adn regenerate enzyme |
| General principles of enzyme catalysis | Geometric effects Nucleophilic attack acid catalysis electrostatic effects molecular recognition water exclusion from hydrophobic cleft |
| Geometric effects | distorted into Half chair makes more reactive and stabilised transition states |
| Nucleophilic attack | enzyme attack reactive group of substrate |
| Acid catalysis | enzyme donate H+ to first product |
| Electrostatic effects | In incorrect mechanism, carbonic ion stabilised by negative group on enzyme |
| Molecular recognition | Diverse and specific recognition of several hexose residues in substrate |
| Competitive inhibitors | Similar shape, bind to AS preventing substrate Vmax unchanged but Km increased (slower rate) Overcome by higher substrate concentration Reversible inhibition e.g. methotrexate chemotherapy drug mimics folic acid, preventing DNA replication |
| Irreversible Inhibition | covalent adduct. much less needed to be potent not recycled targeted against amino acid side chain e.g. sarin in acetylcholinesterase |
| Other inhibitors | Chemically unreactive but use catalytic activity Suicide inhibitor resembles substrate Enzyme reacts to form covalent bond but cannot go further so commits suicide e.g. penicillin with serine in active site of cell wall formation in bacteria |
| Reversible covalent modification | Important and rapid. Mainly eukaryotes Initiated by extracellular signal - hormone/ growth factor e.g. phosphorylation of serine, threonine or tyrosine- catalysed by kinases Serine/ threonine control metabolism, tyrosine control cell growth and differentiation. tyrosine kinase involved in events leading to cancer |
| Genetic Control | Long term control mechanism in higher eukaryotes, short term in bacteria Tryptophan represses production of all enzymes in tryptophan biosynthesis pathway - prevent cell wasting energy on unnecessary production |
| Activation of zymogens | Some enzyme produced and stored in inactive form (zymogen) Can be switched on when needed e.g. blood clotting and digestive enzymes Can only be turned off by destruction |
| Allosteric Enzymes | DO NOT obey Michaelis menten kinetic Often associated with metabolic activity several subunits bind to subunit sigmoidal shaped curve - affinity depends on if any substrate bound already (if bound, easier for next to) |
| Allosteric activators/ inhibitors | Activators shift S curve left (more easily bound) Inhibitors shift S curve right |
| Phosphofructokinase | 4 active sites, used in glycolysis controlled by allosteric effectors inhibited by ATP - feedback inhibition when cell doesn't need energy from glucose enzyme turned off AMP activates - back on when ATP low |
| Cooperative binding | Behave like haem T state has lower oxygen affinity. Converted to higher R affinity by binding When R, easier for rest to bind |
| Compartmentation | Key reaction pathways catalysed by different enzymes Allows greater control so can activate synthesis whilst inhibiting degradation - amplifies change Eukaryotes can do forward and reverse in different compartment, preventing products being immediately used in opposite reaction |
{kind=link}
¿Quieres crear tus propias Fichas gratiscon GoConqr? Más información.