13399155
Descripción
Mapa Mental por veronica velasquez, actualizado hace más de 1 año
|
|
Creado por veronica velasquez
hace más de 6 años
|
|
DISTRIBUCIÓN Y TRANSPORTE DE PROTEÍNAS
- Retículo Endoplásmico (RE)
- Es una red de túbulos y sacos
(cisternas) que se extiende de la
membrana nuclear por todo el
citoplasma. Hay dos tipos
distintos de RE que realizan
diferentes funciones en la
célula, estos son: RE rugoso y RE
de transición
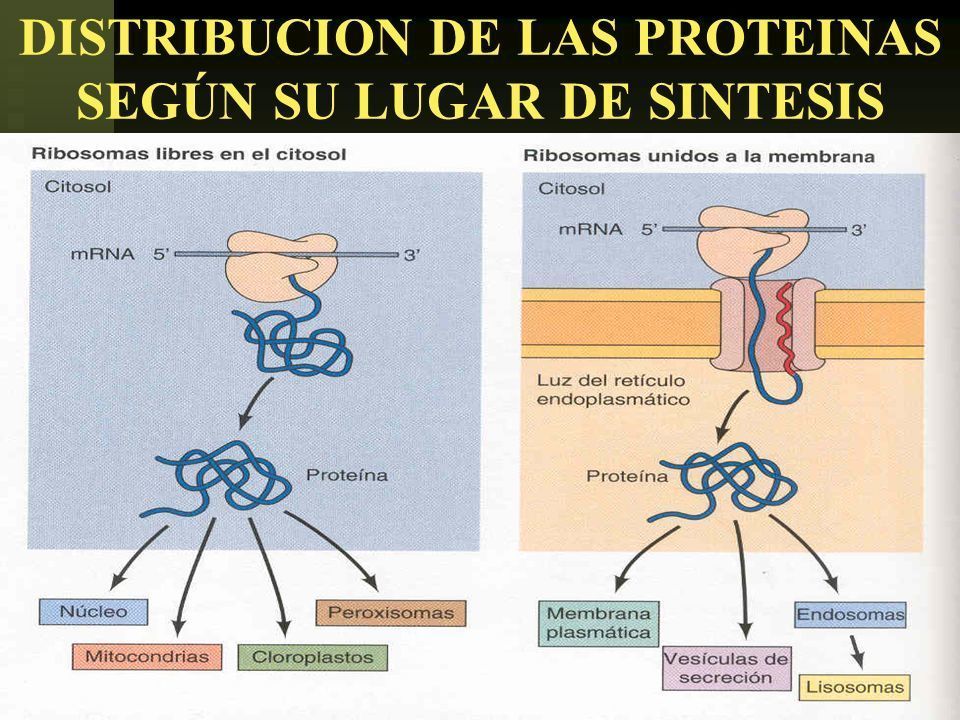
- 1. SECRECIÓN DE PROTEINAS. El
retículo endoplásmico es el primer
cruce de caminos en la distribución de
las proteínas.
- Las proteínas destinadas a la
secreción, a los lisosomas o a la
membrana plasmática se traducen
en los ribosomas unidos a
membrana y son transferidas al RE
rugoso mientras aún se traducen.
- Las proteínas destinadas a la
secreción, a los lisosomas o a la
membrana plasmática se traducen
en los ribosomas unidos a
membrana y son transferidas al RE
rugoso mientras aún se traducen.
- 2. MARCAJE DE LAS PROTEÍNAS PARA DIRIGIRSE AL RE. Las proteínas pueden ser translocadas al RE durante su
síntesis en los ribosomas unidos a la membrana (traslocación cotraduccional) o una vez que la traducción se ha
completado en los ribosomas libres del citosol (Translocación postraduccional)
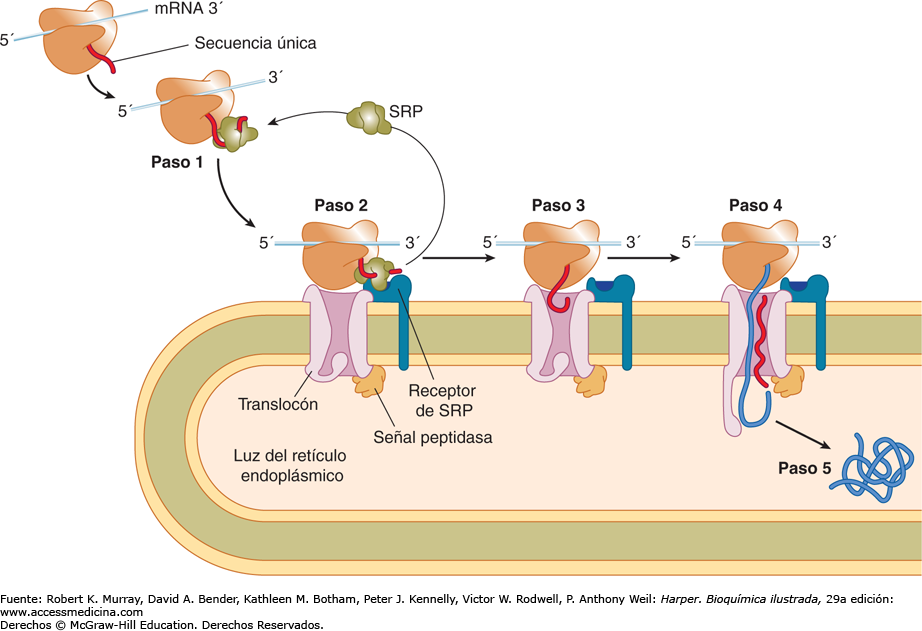
- Translocación cotraduccional. 1) a medida que la secuencia
señal emerge del ribosoma, es reconocida y unida a la
partícula de reconocimiento de la señal (PRS). 2) La PRS
acompaña al complejo hasta la membrana del RE, donde se
une al receptor de la PRS 3) La PRS se libera, el ribosoma se
une a un tranlocon y la secuencia señal se inserta en un canal
de membrana. 4) La secuencia señal abre el translocon, se
reanuda la traducción, y la cadena polipeptídica en
crecimiento es translocada a través de la membrana. 5) La
escisión de la secuencia señal por la peptidasa de la señal
libera el polipéptido en la luz del RE
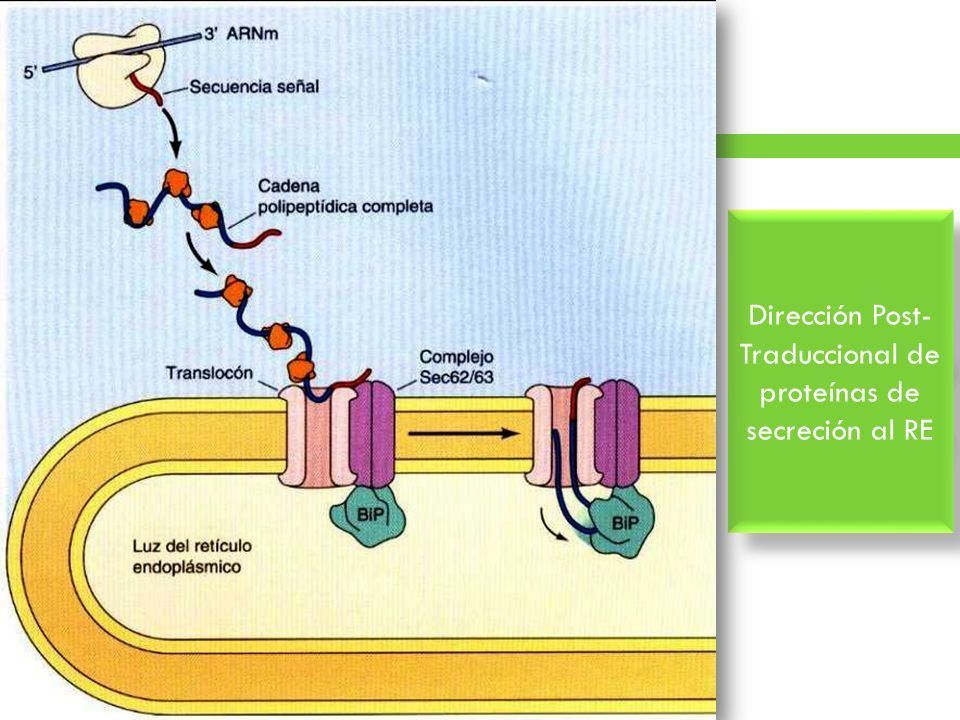
- TRANSLOCACION POSTRADUCCIONAL. Las proteínas destinadas a una internalización
postraduccional se sintetizan en ribosomas libres y se mantienen en una conformación desplegada
mediante chaperonas citosólicas. Sus secuencias señal son reconocidas por el complejo Sec62/63, que
está asociado con el translocon en la membrana del RE. La proteína Sec63 también está asociada con
una proteína chaperona (BiP), que actúa como un "trinquete" molecular dirigiendo la translocación
de la proteína al interior del RE.
- Translocación cotraduccional. 1) a medida que la secuencia
señal emerge del ribosoma, es reconocida y unida a la
partícula de reconocimiento de la señal (PRS). 2) La PRS
acompaña al complejo hasta la membrana del RE, donde se
une al receptor de la PRS 3) La PRS se libera, el ribosoma se
une a un tranlocon y la secuencia señal se inserta en un canal
de membrana. 4) La secuencia señal abre el translocon, se
reanuda la traducción, y la cadena polipeptídica en
crecimiento es translocada a través de la membrana. 5) La
escisión de la secuencia señal por la peptidasa de la señal
libera el polipéptido en la luz del RE
- 3. INSERCIÓN DE PROTEÍNAS EN LA MEMBRANA DEL RE. Las proteínas destinadas a incorporarse en la membrana plasmática o en la del RE,
Golgi o lisosomas se insertan inicialmente en la membrana del RE en lugar de ser liberadas a la luz. Son transportados a lo largo de la ruta RE---->
Golgi----> Membrana plasmática o lisosomas como componente de la membrana, en lugar de como proteínas solubles.
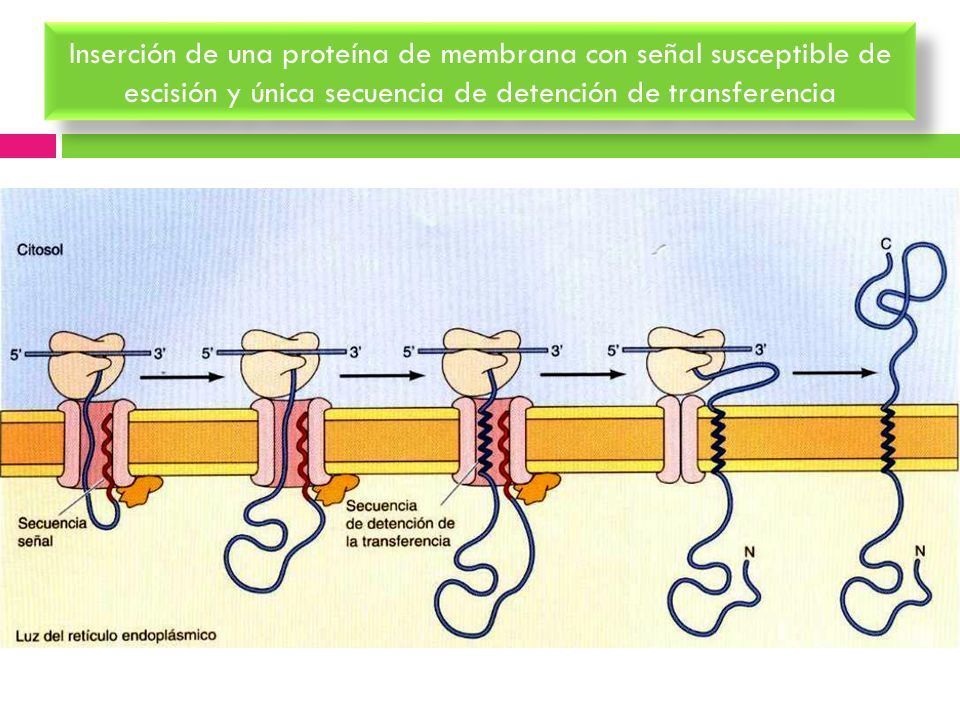
- INSERCIÓN DE UNA PROTEÍNA DE MEMBRANA CON UNA SECUENCIA
SEÑAL SUSCEPTIBLE DE ESCISIÓN Y UNA ÚNICA SECUENCIA DE
DETENCIÓN DE LA TRANSFERENCIA. La secuencia señal se escinde a medida
que la cadena polipetídica atraviesa la membrana, por lo que el extremo amino
terminal de la cadena polipeptídica queda expuesto a la luz del RE. La
translocación es interrumpida cuando el translocon reconoce una secuencia
transmembrana de detención de la transferencia, de moso que el translocon se
cierra y la proteína sale del canal lateralmente y se ancla en la membrana del RE.
El extremo carboxilo teerminal se encuentra en la cara citoplasmática.
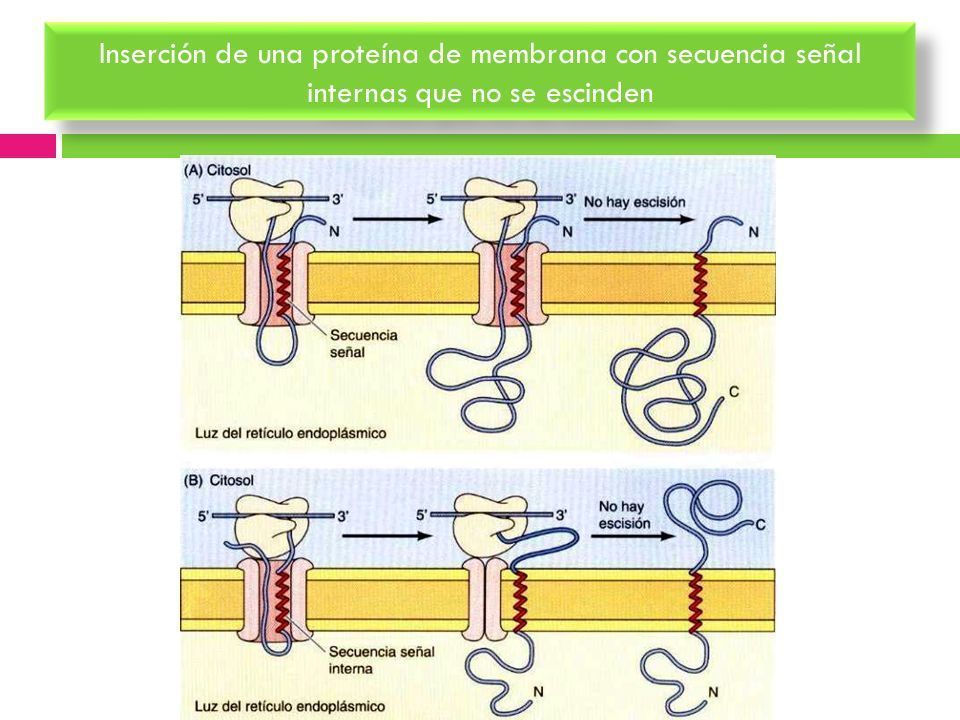
- INSERCIÓN DE PROTEÍNAS DE MEMBRANA CON SECUENCIAS
SEÑAL INTERNAS QUE NO SE ESCINDEN. Las secuencias señal
internas que no se escinden pueden dar lugar a la inserción de cadenas
polipeptídicas en cualquier orientación en la membrana del RE.
- INSERCIÓN DE UNA PROTEÍNA DE MEMBRANA CON UNA SECUENCIA
SEÑAL SUSCEPTIBLE DE ESCISIÓN Y UNA ÚNICA SECUENCIA DE
DETENCIÓN DE LA TRANSFERENCIA. La secuencia señal se escinde a medida
que la cadena polipetídica atraviesa la membrana, por lo que el extremo amino
terminal de la cadena polipeptídica queda expuesto a la luz del RE. La
translocación es interrumpida cuando el translocon reconoce una secuencia
transmembrana de detención de la transferencia, de moso que el translocon se
cierra y la proteína sale del canal lateralmente y se ancla en la membrana del RE.
El extremo carboxilo teerminal se encuentra en la cara citoplasmática.
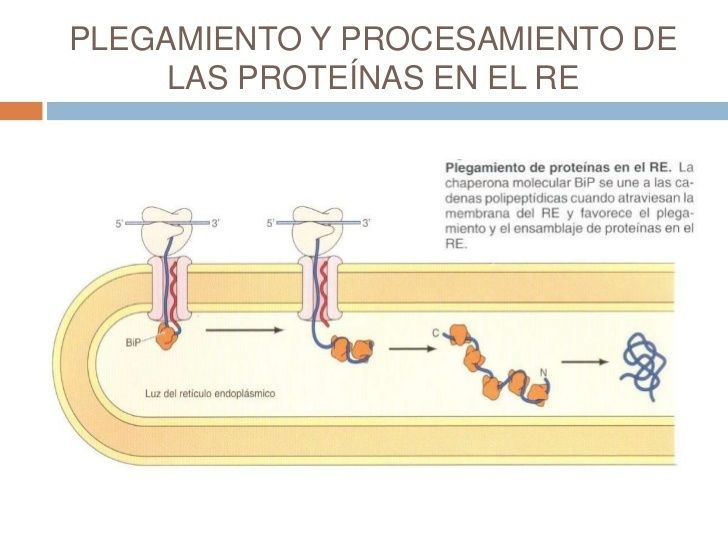
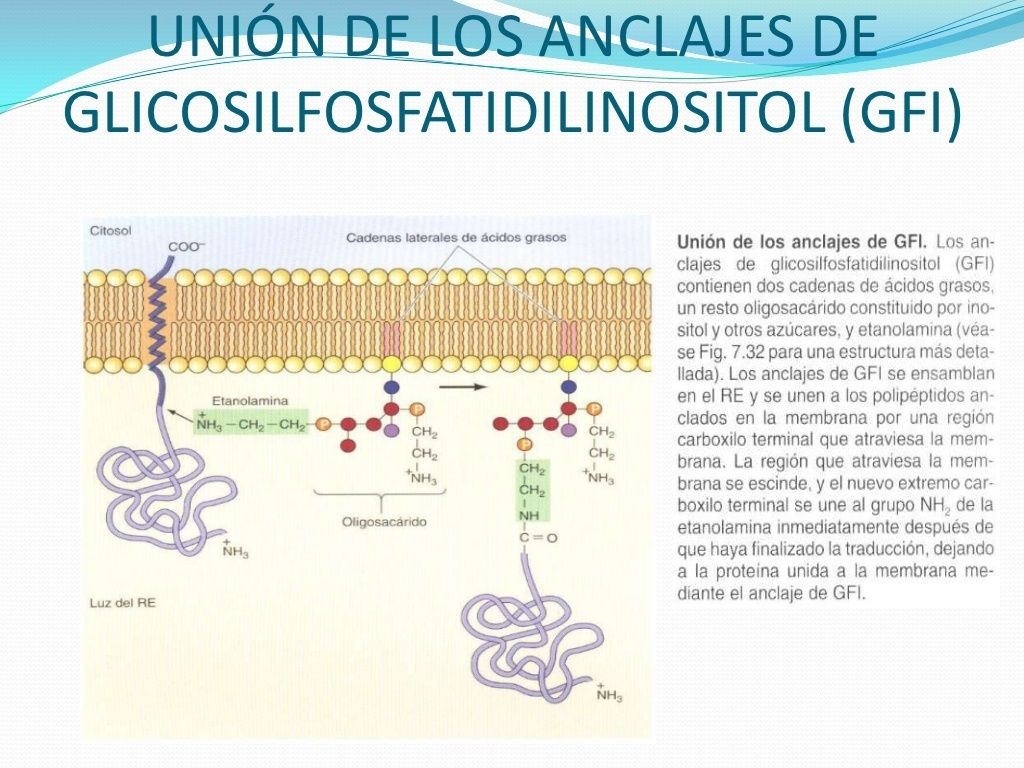
- 4. PLEGAMIENTO Y PROCESAMIENTO DE PROTEÍNAS EN EL RE. Las cadenas polipeptídicas se pliegan en su conformación tridimensional correcta en el RE, asistidos por
chaperonas moleculares que facilitan el plegamiento de las cadenas polipeptídicas. En el RE también tiene lugar la N-glicosilación y la adición de los anclajes GPI.
- La chaperona molecular BiP se une a las cadenas polipeptídicas cuando atraviesan la membrana del REy
favorece el plegamiento y el ensamble de proteínas.
- La chaperona molecular BiP se une a las cadenas polipeptídicas cuando atraviesan la membrana del REy
favorece el plegamiento y el ensamble de proteínas.
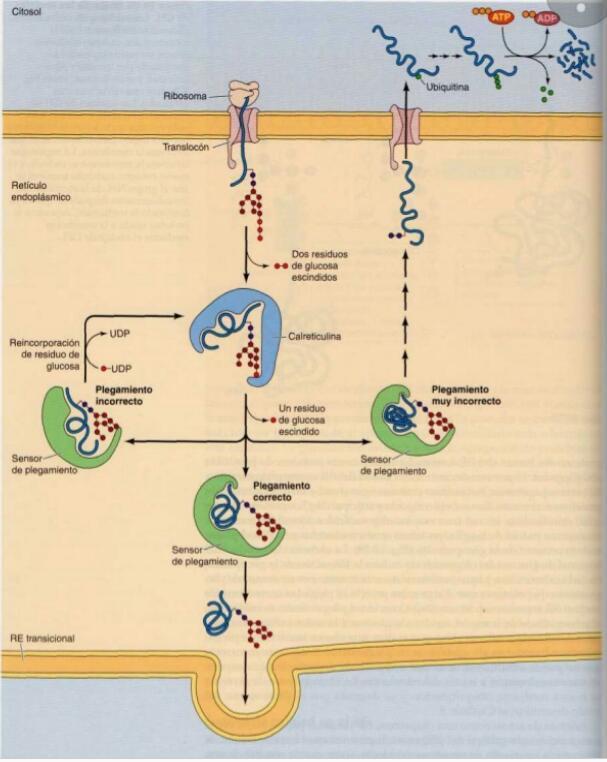
- 5. CONTROL DE CALIDAD EN EL RE. Muchas proteínas secretoras no se pliegan
correctamente la primera vez. Las chaperonas detectan las proteínas plegadas
incorrectamente y las reciclan a través de la vía de plegamiento. Aquellas proteínas
que no pueden ser correctamente plegadas son desviadas de la vía secretora y marcadas
para su degradación.
- PLEGAMIENTO DE PROTEINAS POR
CALRETICULINA.
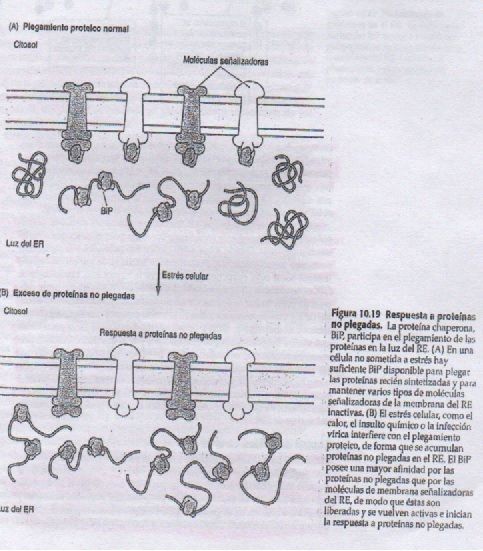
- RESPUESTA A PROTEÍNAS NO
PLEGADAS
- PLEGAMIENTO DE PROTEINAS POR
CALRETICULINA.
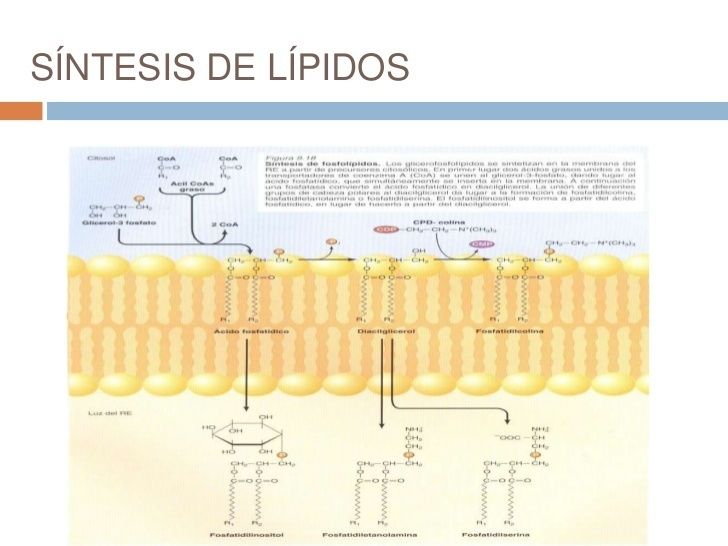
- 6. RE LISO Y SÍNTESIS DE LÍPIDOS. El RE es el sitio principal de
síntesis de lípidos en las células eucariotas, y el RE liso abunda en
las células que tienen un metabolismo lipídico activo y que
realizan la destoxificación de fármacos liposolubles.
- SINTESIS DE FOSFOLIPIDOS
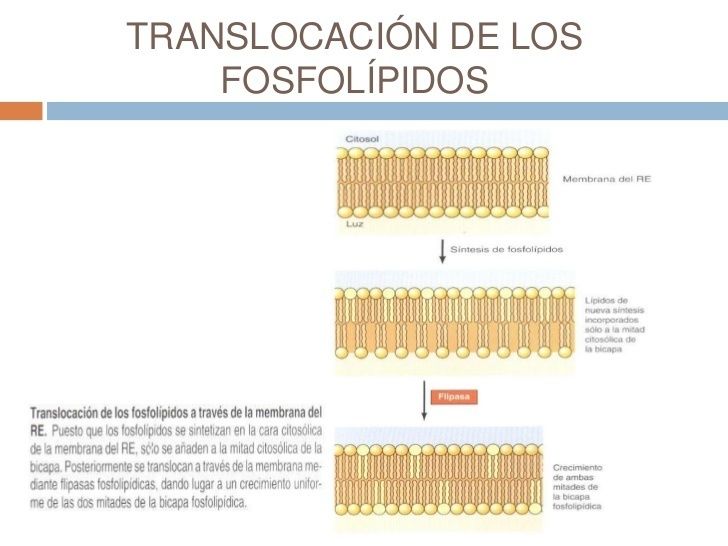
- TRANSLOCACION DE LOS FOSFOLIPIDOS A
TRAVÉS DE LA MEMBRANA DEL RE
- SINTESIS DE FOSFOLIPIDOS
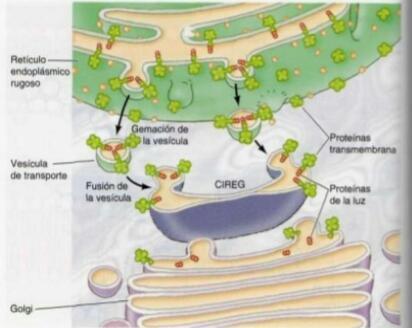
- 7. EXPORTACIÓN DE PROTEÍNAS Y LÍPIDOS DESDE EL RE. Las proteínas y los lípidos
son transportados en vesículas desde el RE al aparato de Golgi. Las secuencias
señalizadoras intervienen en el empaquetamiento selectivo de las proteínas. Las proteínas
residentes en el RE se marcan con otras secuencias señalizadoras que determinan su
regreso al RE desde el Golgi a través de una vía de reciclaje.
- Es una red de túbulos y sacos
(cisternas) que se extiende de la
membrana nuclear por todo el
citoplasma. Hay dos tipos
distintos de RE que realizan
diferentes funciones en la
célula, estos son: RE rugoso y RE
de transición
- APARATO DE GOLGI
- Participa en el
procesamiento y en la
distribución de las
proteínas, así como en
la síntesis de lípidos y
polisacáridos
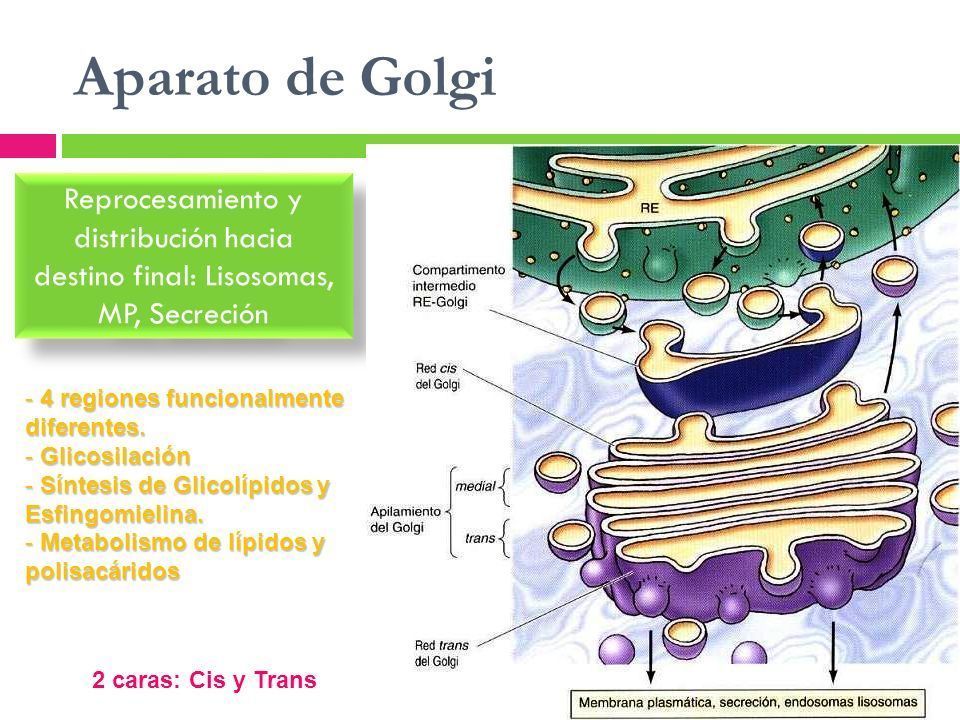
- 1. ORGANIZACIÓN DEL GOLGI.
Morfológicamente está compuesto por
unas bolsas aplanadas, rodeadas de
membrana (cisterna) y por vesículas
asociadas. También está constituido por
cuatro regiones: Red cis del Golgi,
apilamiento del golgi (dividido en
subcompartimientos medial y trans) y la
red trans del golgi.
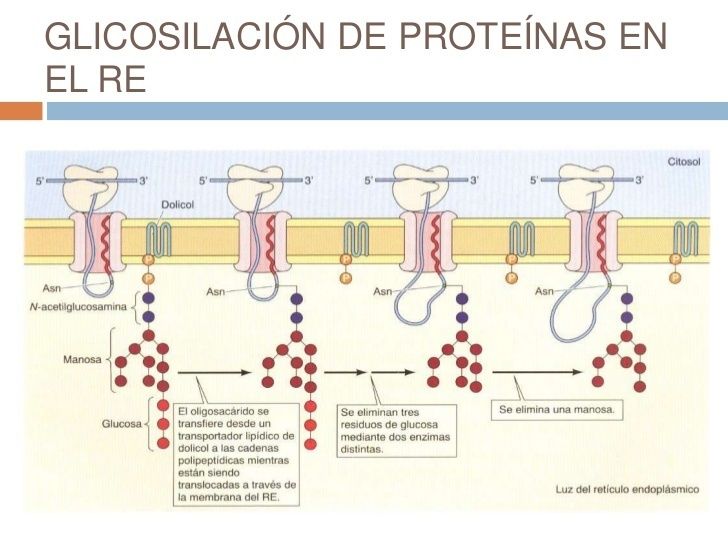
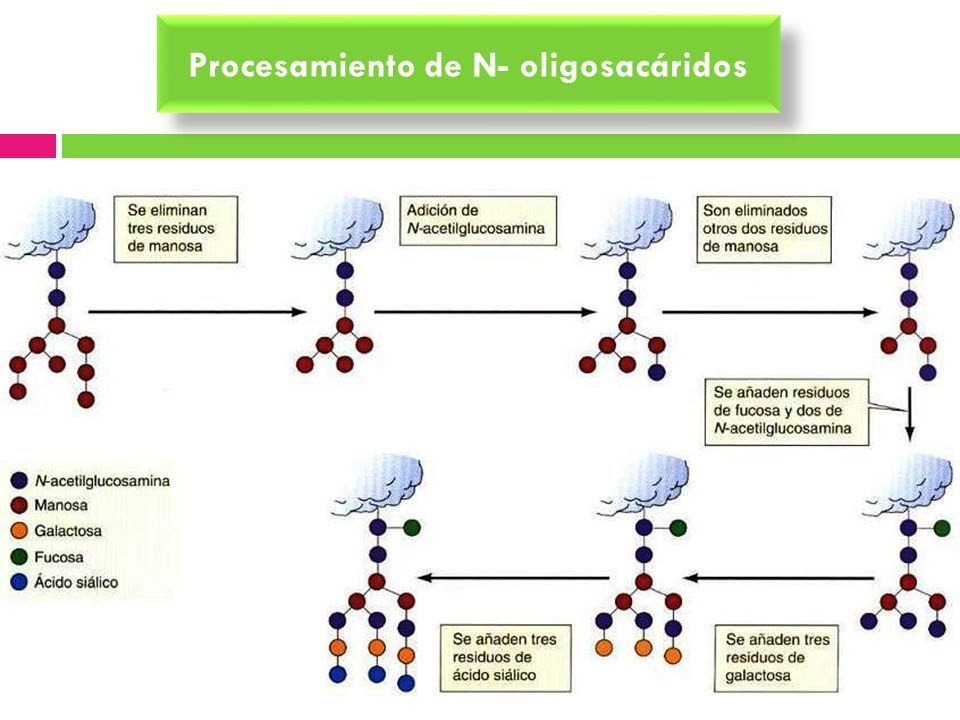
- 2. GLICOLISACIÓN DE PROTEÍNAS EN EL
GOLGI. Los N-oligosacáridos añadidos a las
proteínas en el RE son modificados en el Golgi.
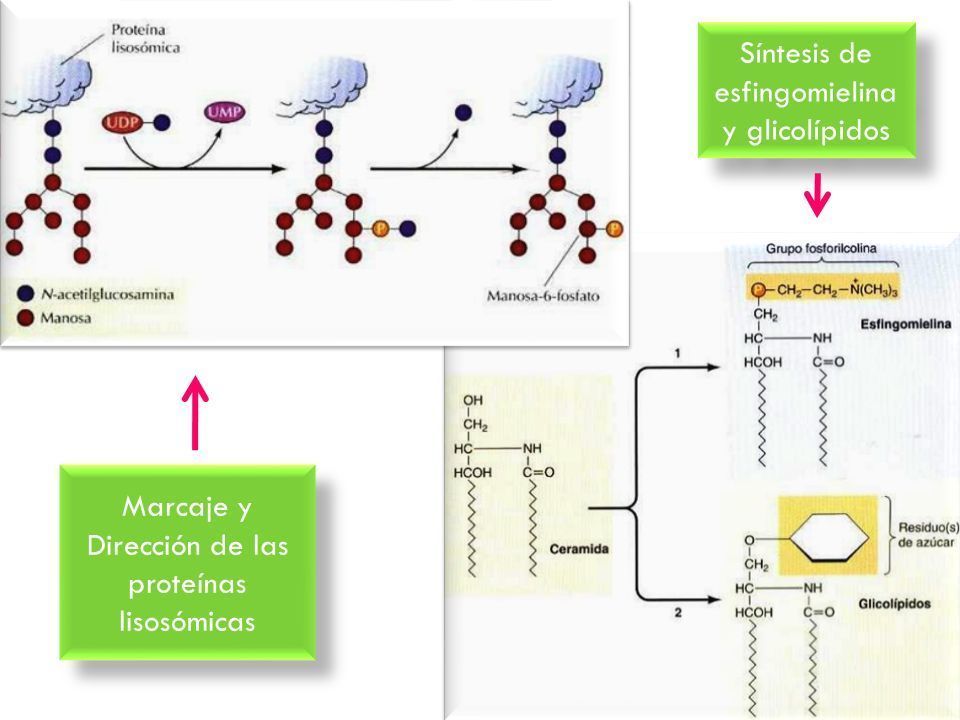
Aquellas proteínas destinadas a los lisosomas son
fosforiladas específiamente en los residuos de
manosa. En el Golgi también tiene lugar la
O-glicosilación.
- 3. METABOLISMO DE LÍPIDOS Y DE
POLISACÁRIDOS EN EL GOLGI. El aparato
de Golgi es el lugar en donde se sintetizan
los glicolípidos, la esfingomielina y los
polisacáridos compuestos de la pared
celular vegetal.
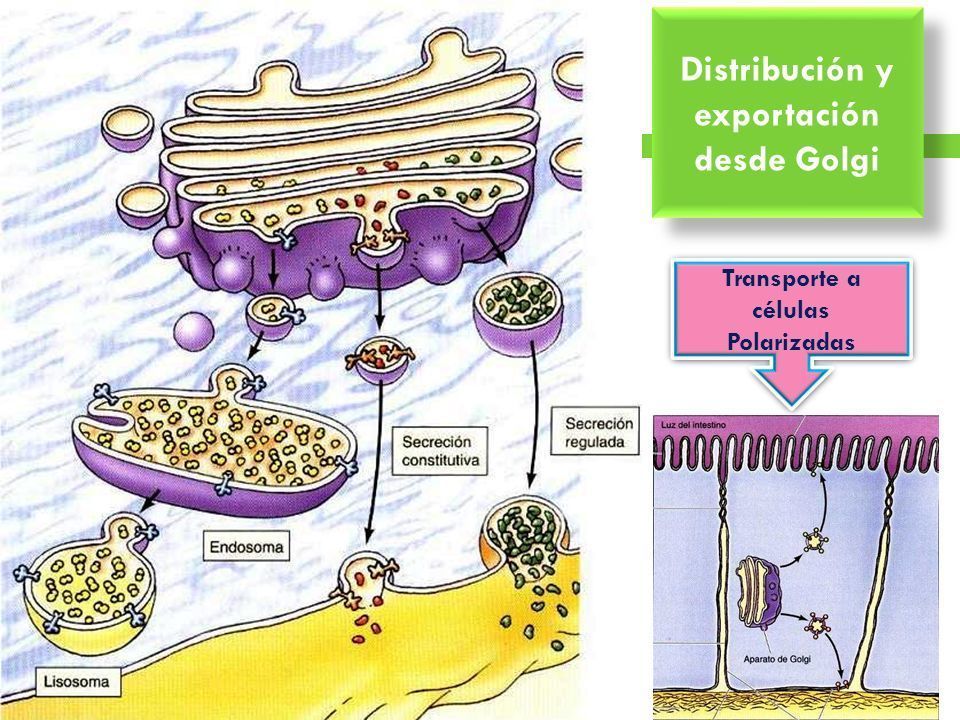
- 4. DISTRIBUCIÓN Y EXPORTACIÓN DE
PROTEÍNAS DESDE EL APARATO DE GOLGI.
La vía secretora constitutiva, que está presente en
todas las células, da lugar a una secreción de
proteínas continua no regulada. Algunas células
también poseen una vía secretora regulada
diferente, a través de la que secretan proteínas
específicas en respuesta a señales ambientales.
- Las proteínas se distribuyen en la red trans del Golgi para
ser empaquetadas en vesículas de transporte que se dirigen
a ser secretadas, a la membrana plasmática, a los lisosomas
o a las vacuolas de las levaduras y plantas.
- Las proteínas se distribuyen en la red trans del Golgi para
ser empaquetadas en vesículas de transporte que se dirigen
a ser secretadas, a la membrana plasmática, a los lisosomas
o a las vacuolas de las levaduras y plantas.
- Participa en el
procesamiento y en la
distribución de las
proteínas, así como en
la síntesis de lípidos y
polisacáridos
- MECANISMO DE TRANSPORTE DE LAS
VESÍCULAS
- El transporte de las vesículas es una actividad celular fundamental,
responsable del tráfico molecular entre diversos compartimientos
rodeados por membrana específicos. Por lo tanto, la selectividad de
dicho transporte resulta clave para mantener la organización
funcional de la célula.
- 1. APROXIMACIONES EXPERIMENTALES AL CONOCIMIENTO DEL
TRANSPORTE DE LAS VESÍCULAS. El mecanismo de transporte de las
vesículas se ha determinado mediante el estudio de mutantes de levaduras,
de sistemas reconstruidos de células libres y de vesículas sinápticas,
visualización del tráfico de proteínas a través de la vía secretora y análisis
proteómico.
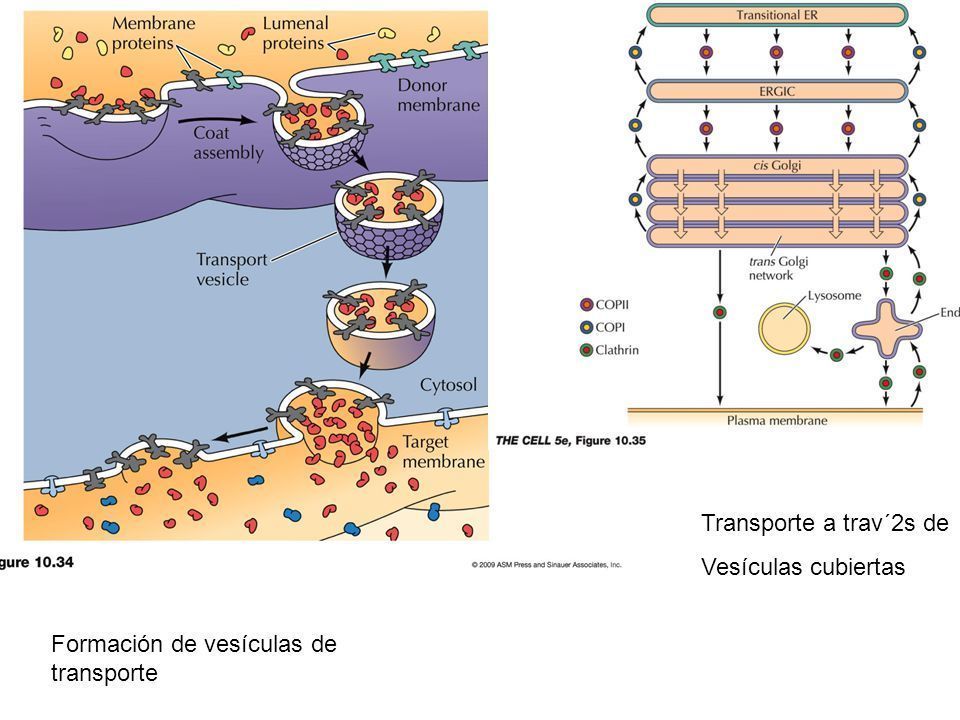
- 2. SELECCIÓN DE LA MERCANCÍA, PROTEÍNAS DE
REVESTIMIENTO Y GEMACIÓN CELULAR. Las
vesículas de transporten que llevan proteínas
secretoras desde el RE a otros compartimientos
posteriores están recubiertas con proteínas de la
cubierta citosólica, y por tanto se denominan vesículas
cubiertas.
- Se conocen tres familias de proteínas de
cubierta de las vesículas: Clatrina,
transporta su mercancía en sentido
retrógado desde la red trans del Golgi; la
COPI, se encarga de recuperar proteínas
residentes del CIREG y la red cis del Golgi;
la COPII transportan su mercancía del RE al
aparato de Golgi.
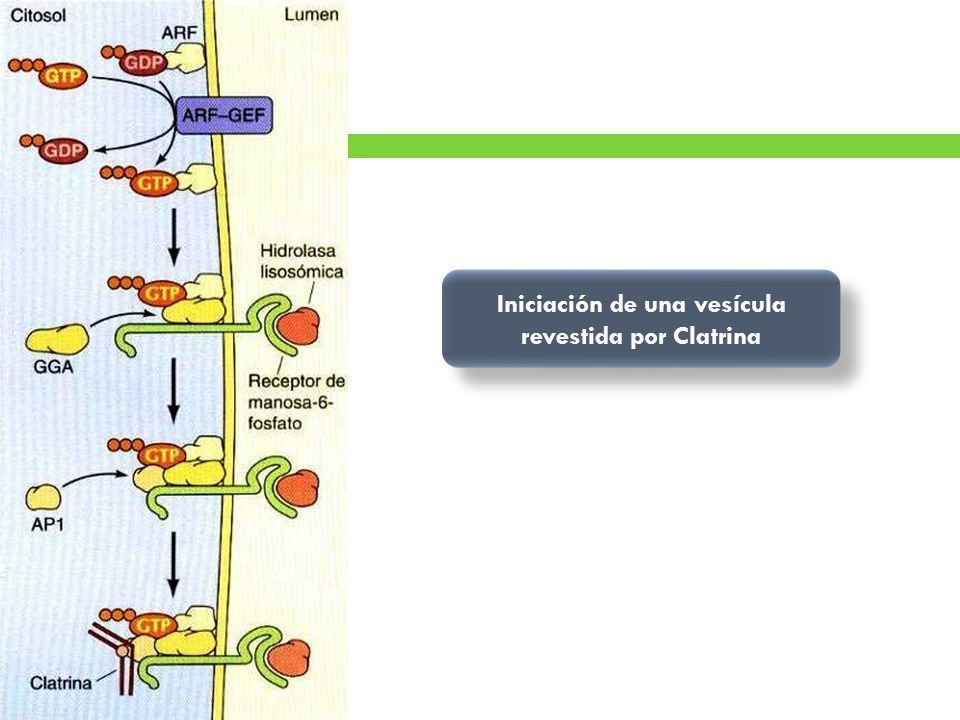
- Las moléculas específicas que se transportan se
seleccionan por medio de complejos de pequeñas
proteínas de unión a GTP (en este caso la ARF) y
proteínas adaptadoras que se asocian a las proteínas de
cubierta.
- Se conocen tres familias de proteínas de
cubierta de las vesículas: Clatrina,
transporta su mercancía en sentido
retrógado desde la red trans del Golgi; la
COPI, se encarga de recuperar proteínas
residentes del CIREG y la red cis del Golgi;
la COPII transportan su mercancía del RE al
aparato de Golgi.
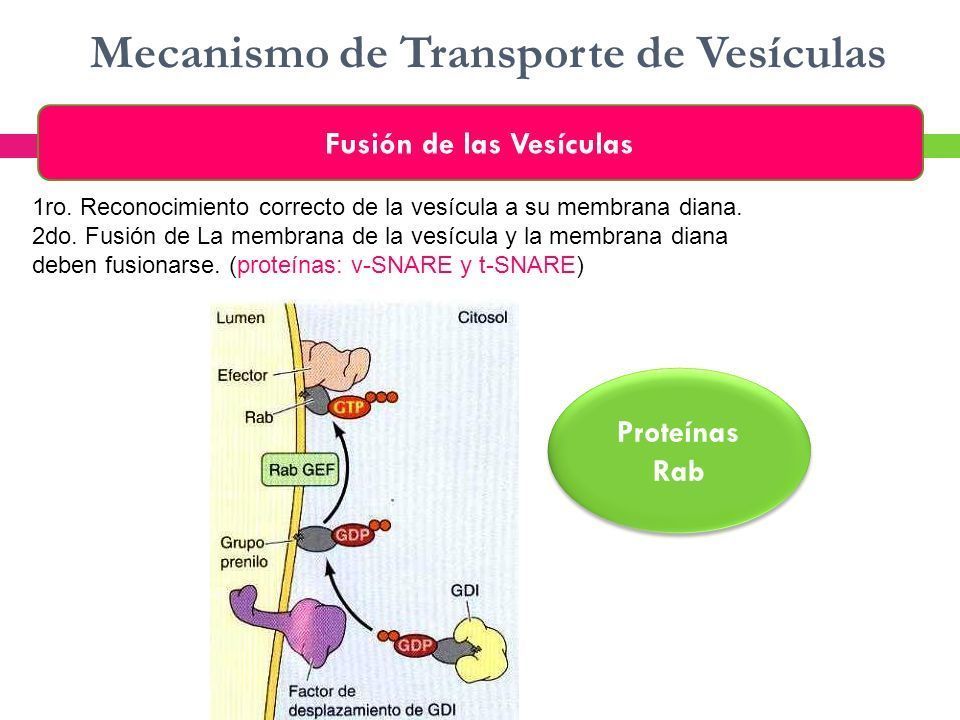
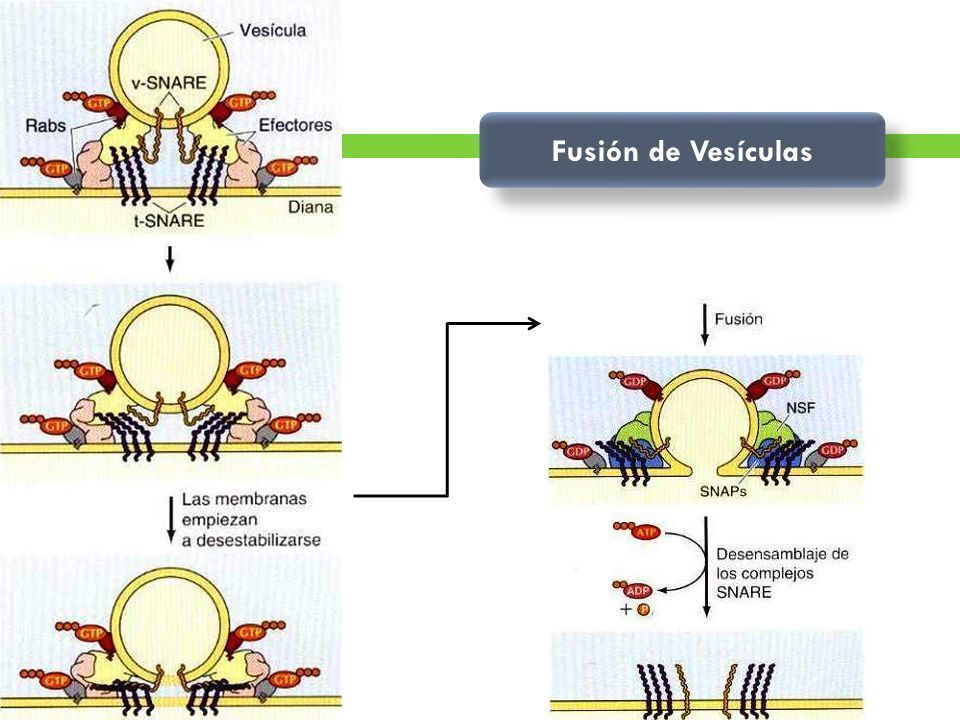
- 3. FUSIÓN DE LAS VESÍCULAS. La unión de las vesículas con la membrana diana
adecuada está mediada por interacciones entre pares de proteínas transmembrana
(las SNARE, y las efectoras) y pequeñas proteínas de unión a GTP (el Rab), lo que da
lugar a la fusión de las membranas. algunos tipos de fusión con la membrana
plasmática (exocitosis) tienen lugar en complejos multiproteicos específicos
denominados exocistos.
- El transporte de las vesículas es una actividad celular fundamental,
responsable del tráfico molecular entre diversos compartimientos
rodeados por membrana específicos. Por lo tanto, la selectividad de
dicho transporte resulta clave para mantener la organización
funcional de la célula.
- LISOSOMAS
- Los lisosomas son organulos rodeados de membrana que
contienen una serie de enzimas capaces de degradar todas las clases
de polímeros biológicos (proteínas, ácidos nucleicos, glúcidos y
lípidos). Los lisosomas funcionan como el sistema digestivo de
la célula, sirviendo tanto para degradar el material capatado del
exterior de la célula como para digerir los componentes
obsoletos de la propia célula.
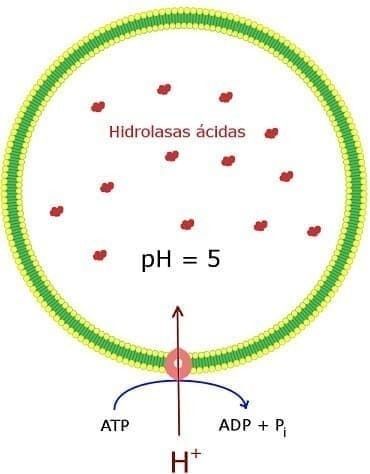
- 1. HIDROLASAS ÁCIDAS LISOSÓMICAS. Los lisosomas contienen una serie de hidrolasas
ácidas que degradan proteínas, ácidos nucleicos, polisacáridos y lípidos. Estas enzimas son
activas al pH ácido (aproximadamente 5) del interior de los lisosomas pero no al pH neutro
(aproximadamente 7,2) del resto del citoplasma. Para mantener su pH ácido interno, los
lisosomas bombardean la membrana lisosómica para atraer iones H+ desde el citosol.
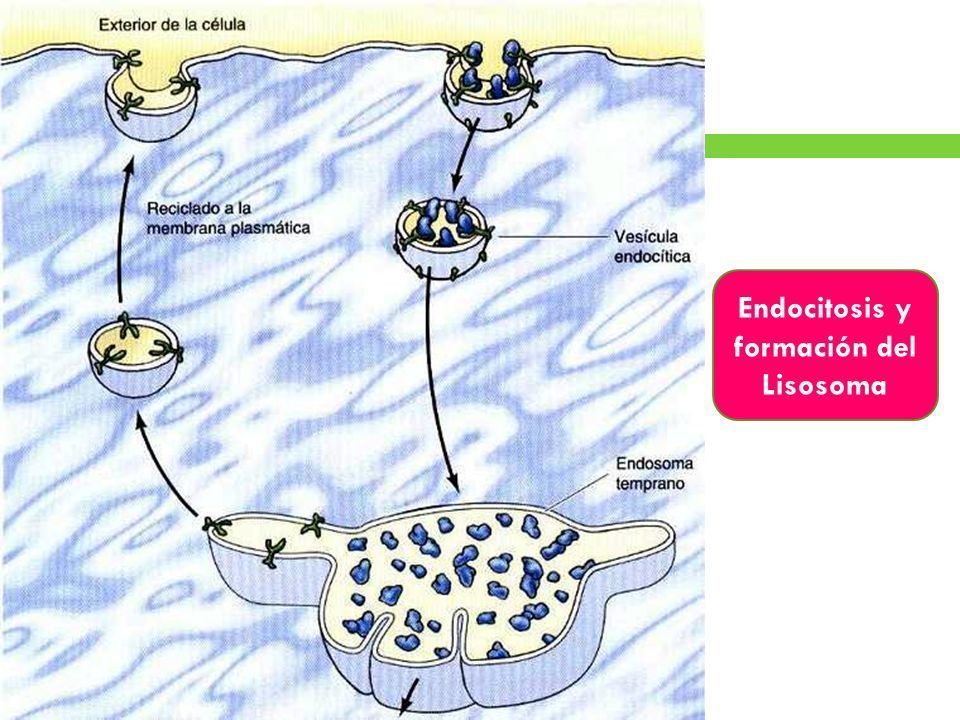
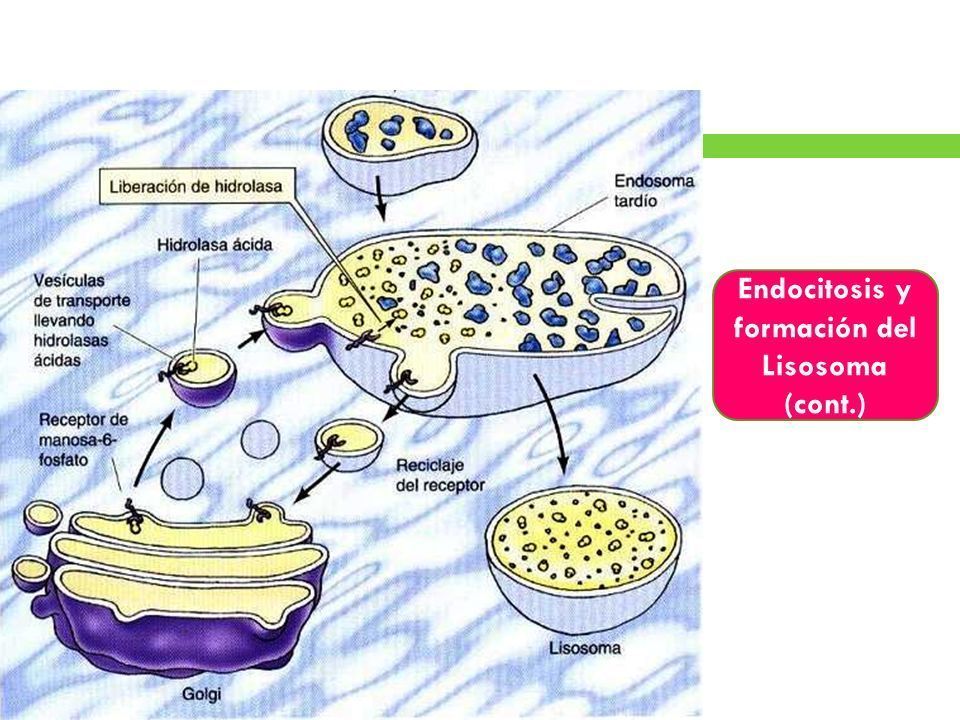
- 2. ENDOCITOSIS Y FORMACIÓN DE LOS LISOSOMAS. La formación de los endosomas y de los
lisosomas representa una intersección entre la vía secretora, mediante la que se procesan las
proteínas lisosómicas, y la vía endocítica, mediante la cual son ingeridas las moléculas
extracelulares a partir de la superficie celular. Estas moléculas son transportadas a los
endosomas tempranos, que pueden madurar para formar lisosomas a medida que se traen
hidrolasas ácidas desde el Golgi.
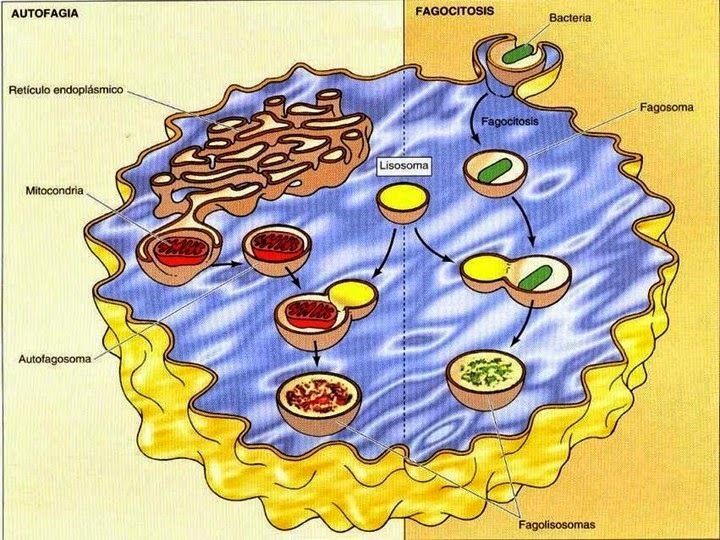
- 3. FAGOCITOSIS Y AUTOFAGIA. Los lisosomas son
responsables de la degradación de partículas grandes, ingeridas
mediante la fagocitosis, y de la digestión gradual de los propios
componentes de la célula mediante la autofagia (también
desarrolla un papel importante en la muerte celular programada)
- Los lisosomas son organulos rodeados de membrana que
contienen una serie de enzimas capaces de degradar todas las clases
de polímeros biológicos (proteínas, ácidos nucleicos, glúcidos y
lípidos). Los lisosomas funcionan como el sistema digestivo de
la célula, sirviendo tanto para degradar el material capatado del
exterior de la célula como para digerir los componentes
obsoletos de la propia célula.
Recursos multimedia adjuntos
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
¿Quieres crear tus propios Mapas Mentales gratis con GoConqr? Más información.