Description
|
|
Created by Joakim Silva
almost 9 years ago
|
|
Page 1
Objectives:- To know that specificity can be achieved through the formation of signalling complexes.- To know that the modular architecture of signalling proteins allows for cooperative interaction.- To know that membrane confinement facilitates intermolecular interactions.- To know that the duration and speed of a response to a signal can range from milliseconds to days.- To know that positive feedback responses are used to generate a switch-like (all or nothing) response.- To know that negative feedback responses are used to generate adaptive responses and oscillations. - To know that a combination of feedback loops with logic gates can generate complex response behaviour. Lecture 2: Explains how cell signalling specificity is achieved in a noisy cellular environment. This is achieved using multiprotein complexes, modular architecture of signalling proteins, signalling networks and feedback loops.
Page 2
Signalling Complexes
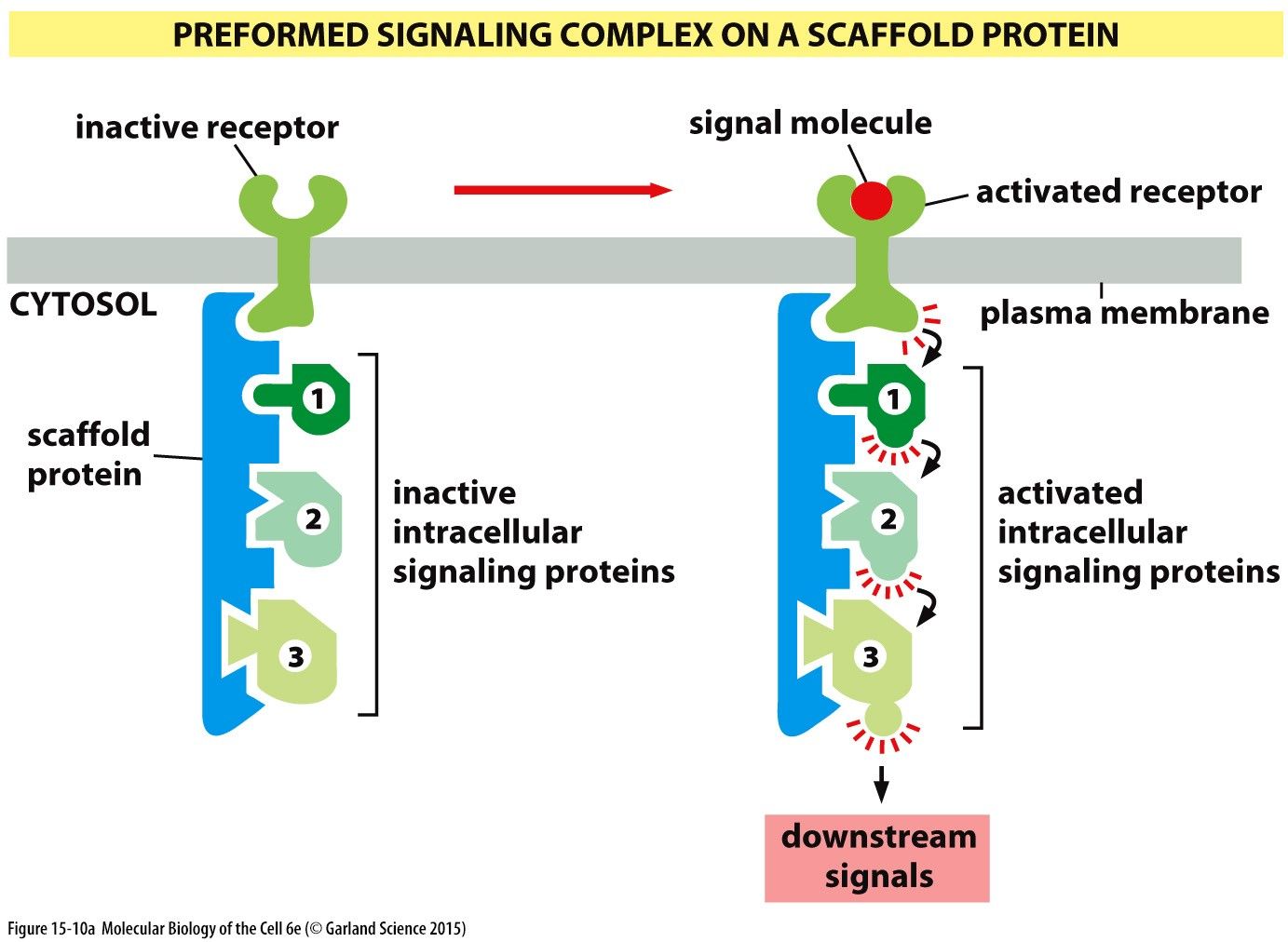
Signalling Complex I: Preformed signalling complex on a scaffold proteinSignalling complex's on a scaffold protein are comprised of a receptor protein, a scaffold protein and a number of inactive intracellular signalling proteins. The scaffold protein is bound to the cytosolic side of the receptor and has binding regions for the intracellular signalling proteins. When a signalling molecule binds to the receptor it is activated. This causes a structural change in the receptor that activates the intracellular signalling proteins bound to the scaffold protein activating downstream signal pathways.
{kind=link}
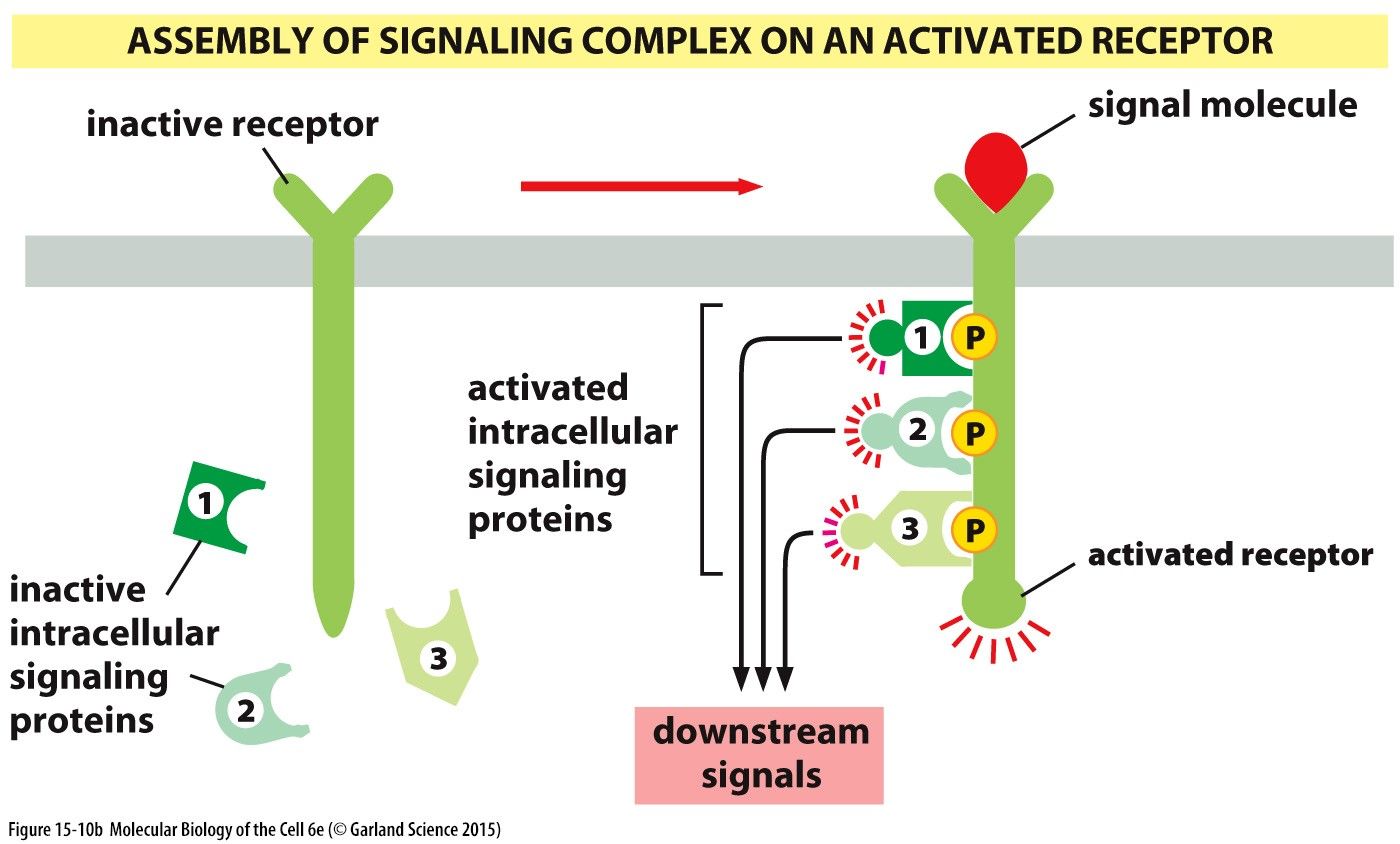
Signalling Complex II: Assembly of signalling complexes on an activated receptorAn inactive receptor embedded in the phospholipid bilayer is activated by a signal molecule on the extracellular side. This activates the receptor such that inactive intracellular signalling proteins may now dock and its docking sites on the active receptors cytosolic side. This in turn activates the intracellular signalling proteins triggering downstream signalling pathways.
{kind=link}
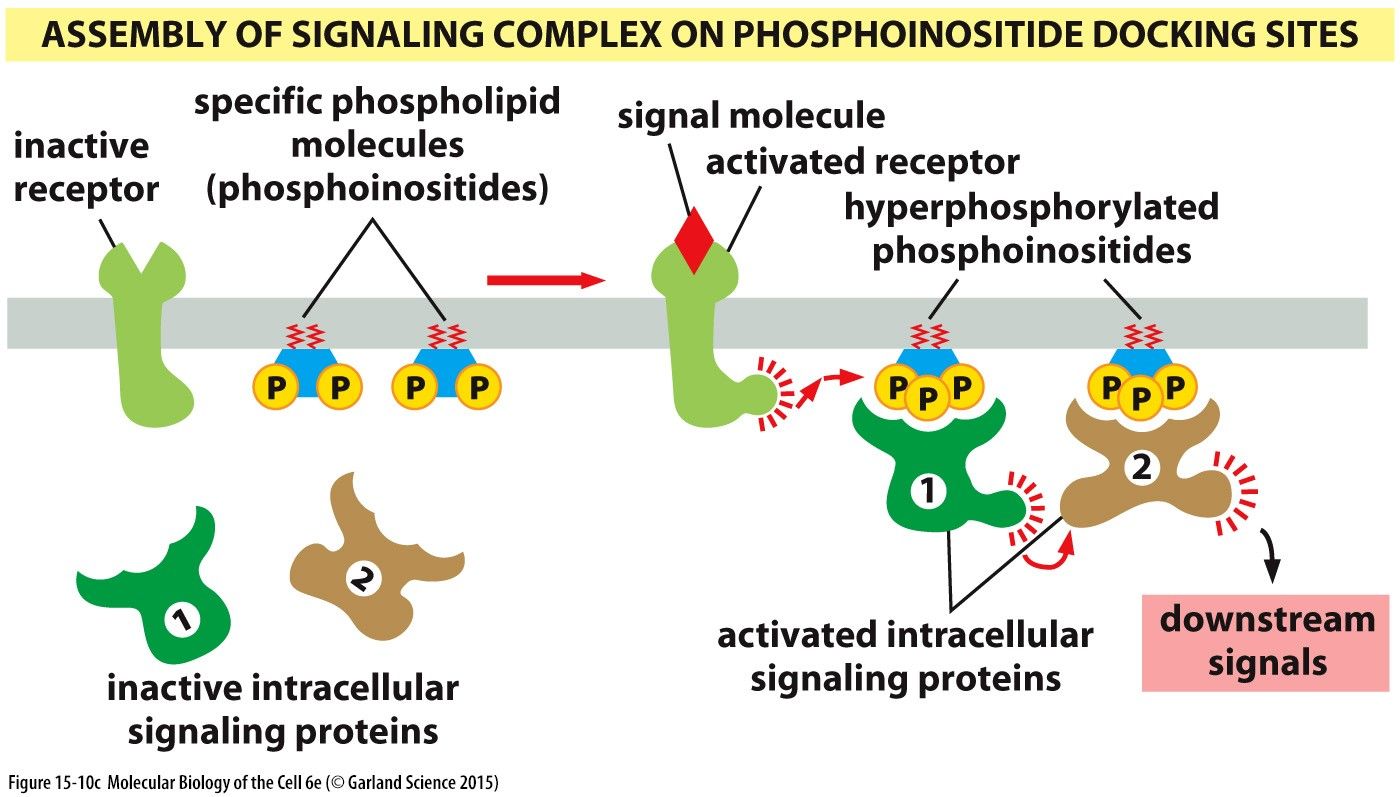
Signalling Complexes III: Assembly of signalling complex on phosphoinositide docking sitesComprised of an inactive receptor and specific phospholipid molecules (phosphoinositides) embedded in the phospholipid bilayer. When a signal molecule binds to the receptor it becomes active causing the hyperphosphorylation of the phosphoinositides to their hyperphosphorylated state. When intracellular signalling proteins dock at these hyperphosphorylated phosphoinositide sites they are activated triggering downstream signal pathways.
{kind=link}
Page 3
Modular architecture of signalling allows for cooperative recognition: Different combinations of domains have their own binding patterns and activity. Linking domains results in a higher affinity and increases specificity for binding, therefore binding may occur at low concentrations for receptors, proteins and intracellular signalling proteins.A high density of binding sites favours rapid rebinding: A high density of receptor tail phosphorylation increases the likelihood of intracellular signal protein of finding a binding partner, and the chance of rebinding. However a low density of receptor tail phosphorylation would mean once an intracellular signal protein dissociates it is likely to be lost. Another example is phosphoinositol lipids, whereby clustering these receptors results in strong binding and spreading sparsely results in weak binding.Membrane association makes molecular interactions more efficient: By anchoring a structure that would otherwise have 6 degrees of freedom, there is only one way that two molecular interactions may come together. Therefore anchoring means a stronger binding affinity.
Signalling Systems: Persistence & Sensitivity of Response:Transient Response - Fast turnover of signal mediators, negative feedback loops, adaption.Permanent Response - Switch-like behaviour, positive feedback loopsLow Sensitivity - A linear response as concentration increases.Ultrasensitive - Cooperativity (allosteric regulation) gives a sigmoidal response to increase in concentration. Once a threshold is reached we see a positive feedback loop for a period of time until binding sites become saturated.
{kind=link}
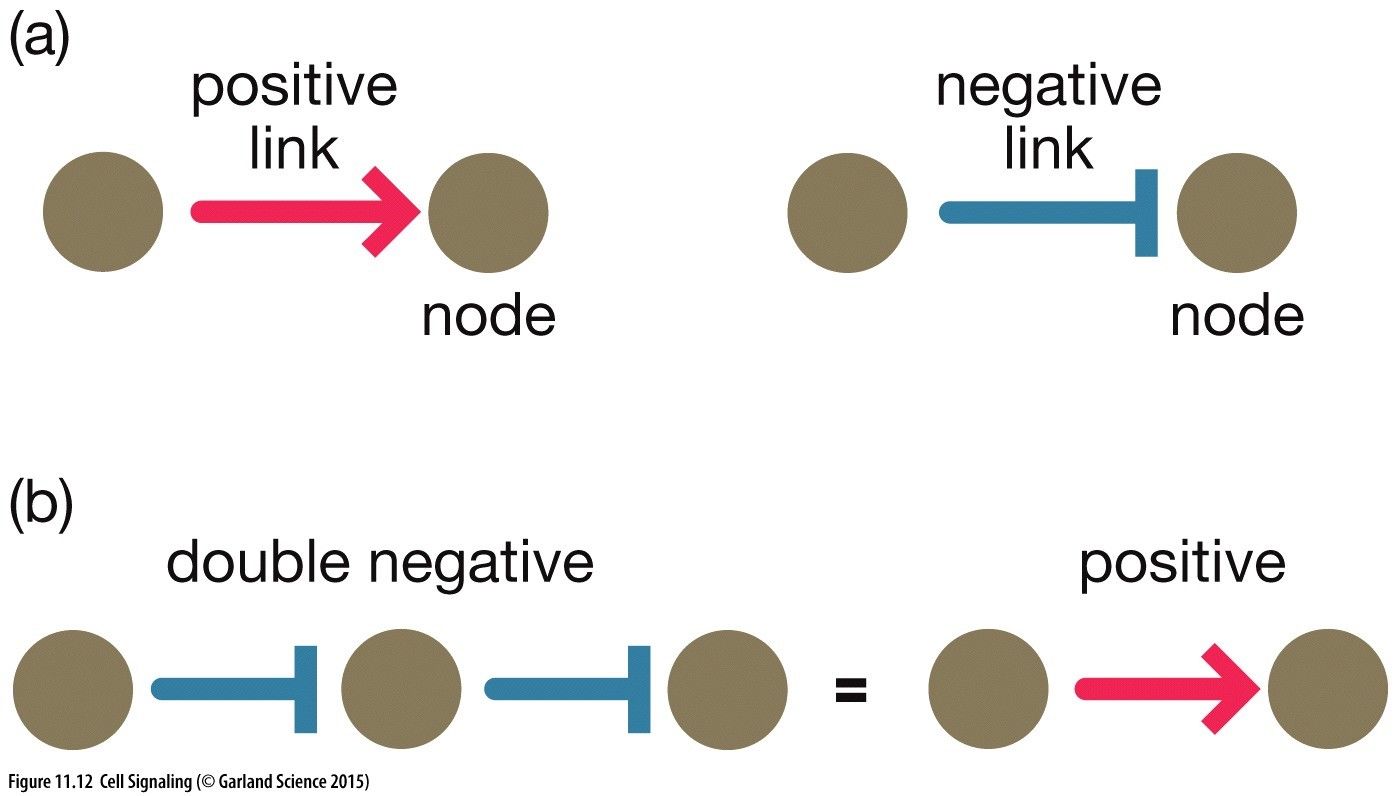
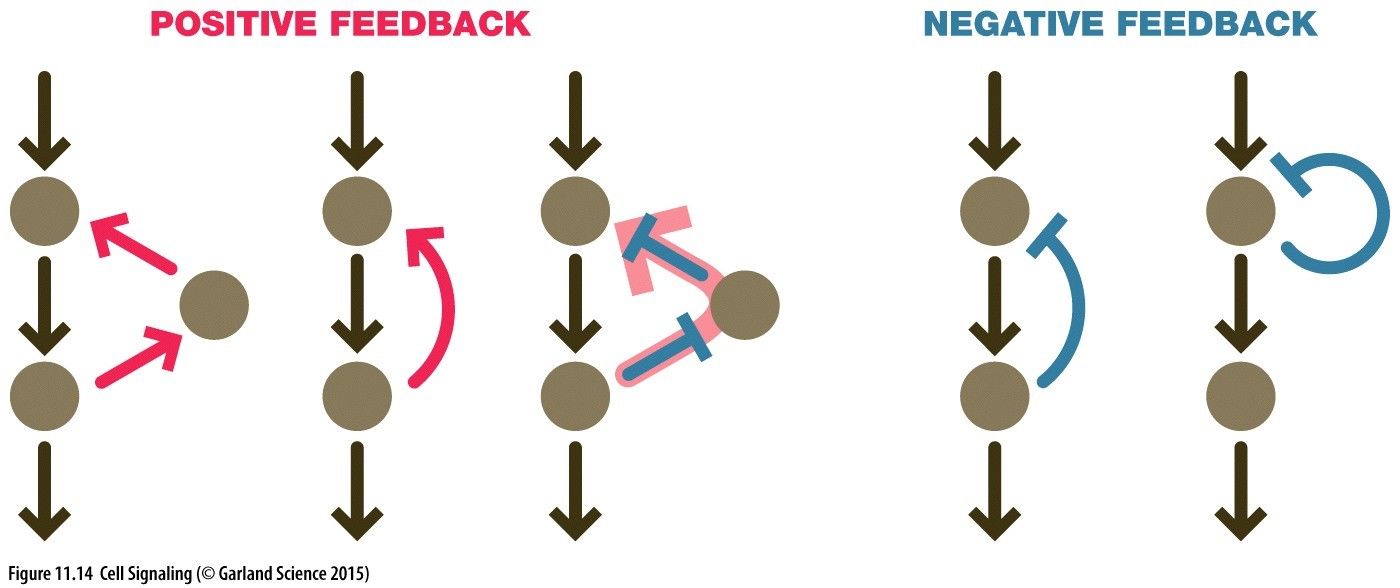
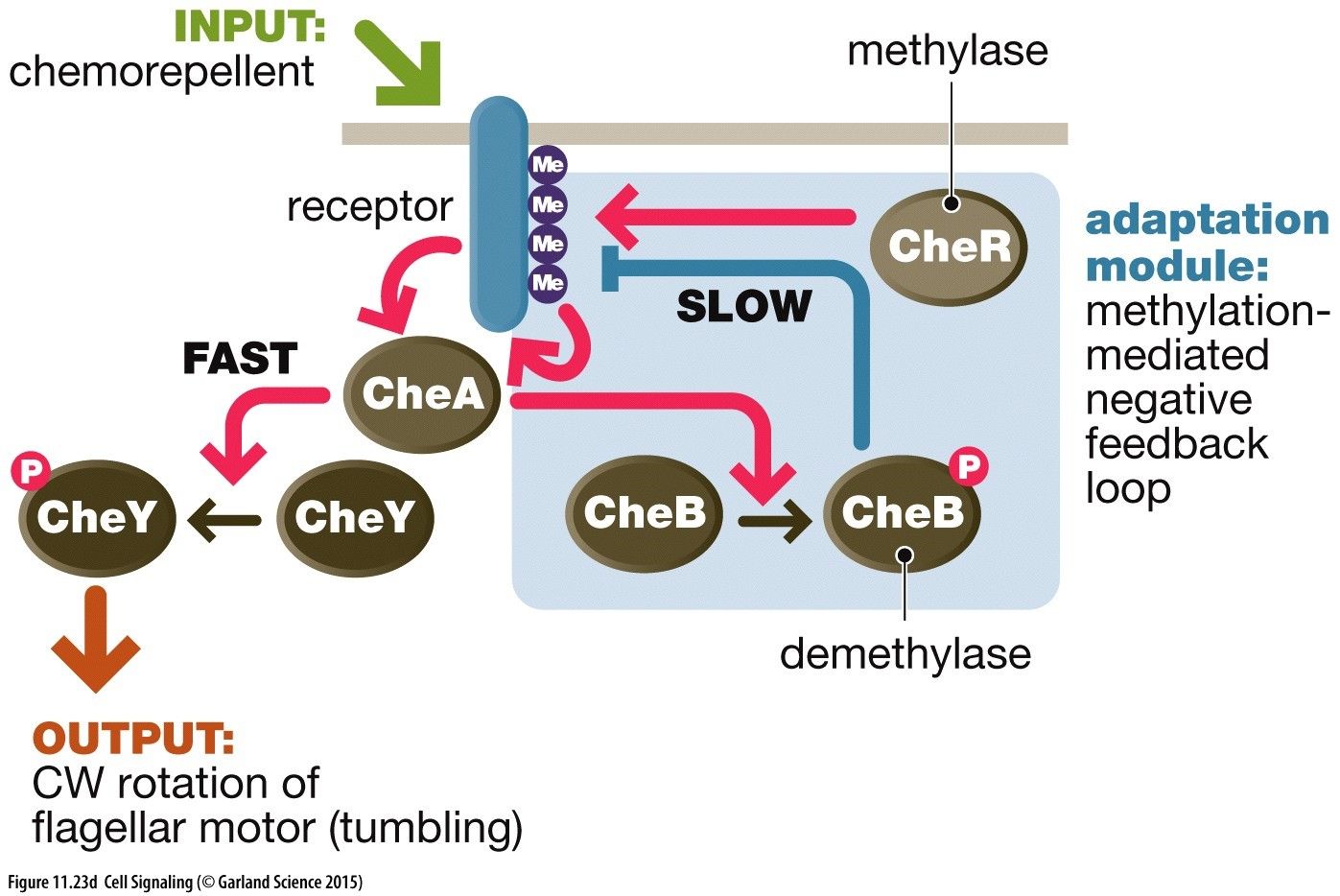
Signalling Network Notation(a) Positive Link = A activates node B Negative Link = C inhibits D(b) Double Negative = E inhibits F, preventing inhibition of F on G = Positive Amplification of GPositive Feedback Loops: Amplification of a pathway deviating away from the norm. Node I positively induces node II to achieve an output. Node II also increases further positive signalling of node III. Node III positive induces node I causing a positive feedback loop. Node I positive induces node II to achieve an output. Node II also increases further signalling of node I causing a positive feedback loop. Node I positively induces node II to achieve an output. Node II inhibits node III which is itself inhibiting node I. This causing a positive signalling of node I leading to a positive feedback loop. Amplification in signalling cascades: We can achieve an amplification of a signal involving a small input via signalling cascades. For instance an input activates kinase I, that go on to phosphorylate other molecules and kinase II, that then activate an even larger number of kinase III amplifying this signal to achieve an output. However, this does need to be tightly regulated with feedback loops to coordinate inputs and outputs of cell behaviour. This is often allosterically regulated and occurs in a switch like manner.Negative Feedback Loop: Regulation of a signalling pathway to return signal back to the norm. Node I activates node II to achieve an intended output. Node II directly inhibits node I maintaining signalling pathway to narrow bounds. Node I activates node II to achieve an intended output. Node II is an inhibitor of itself and will therefore inhibit the signalling pathway if the concentration of node I is high. Adaption: (Bacteria Examples) A chemorepellant input binds to a receptor, CheR methylase methylates the receptor. This activates kinase CheA to phosophorylate CheY --> Activated CheY-Pi which rotates the flagellum motor for tumbling. This then needs to be turned off after a while which is achieved with CheB demethylase. In addition to rotating the flagellum a slower pathway with a bit of delay CheA also phosphorylates CheB which then demethylates the receptor turning off the rotating motor.
{kind=link}
{kind=link}
Page 4
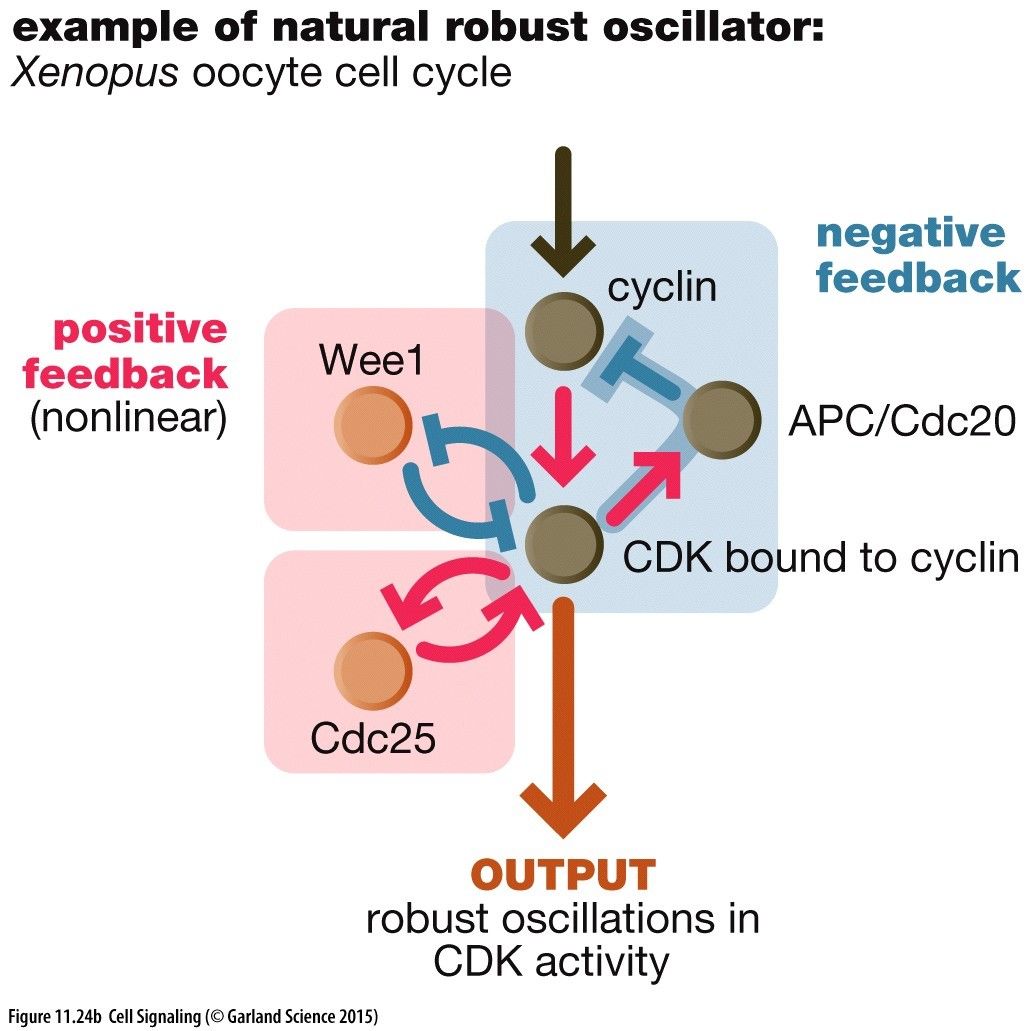
Other Adaption Mechanisms: Another way is a system where the signal protein that binds to the receptor is removed turning off the pathway.Negative Feedback Oscillations: If the negative feedback mechanism is delayed we get oscillations of proteins in the pathway. This will tend towards a stable oscillation.e.g. Xenopus oocyte cell cycleCyclin turns on the cyclin dependent kinase (CDK) to bring about an output within the cell. There is a negative feedback loop wherebye CDK activates APC that unuiquitinates the cyclin and sends it off for destruction. This turns off what is upstread of the cyclin.There is also a positive feedback by a kinase Wee1 that inhibits CDK by phosphorylating it. CDK phosophorylates Wee1 inhibiting it also.There is two positive feedback loops of Cdc25 to CDK also giving a robust oscillation of this mechanism.
{kind=link}
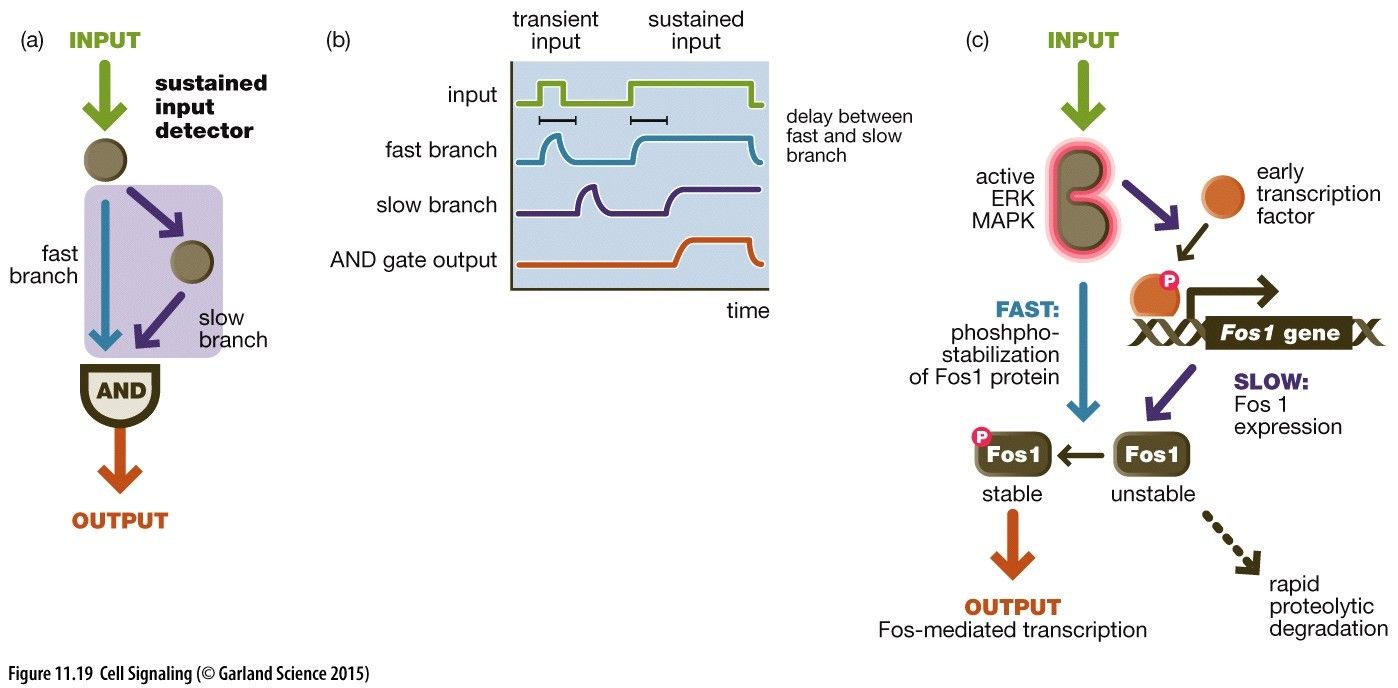
Logic Gates: This is where a pathway only occurs when two separate molecule inputs A + B are present at the same time (AND Gate) to get an output.Therefore there are two binding regions on a protein complex, both signalling molecules a and b must bind to receptor A and B to see an activation of pathway at this AND gate to achieve an output.Coherent Feed Forward Loop: For a sustained input Where two branches converge on an AND gate. This is both a SLOW and a FAST branch. Therefore only when the SLOW branch signal reaches with a sustained input will a combination of the two cause as output. If the SLOW branch is not sustained then by the time this reaches the AND gate the FAST branch would have already reached with a diminished signal giving no output.e.g.Fos mediated transcription via an ERK/MAPK phosphate cascade: Input activates ERK/MAPK whose FAST branch stabilises unstable Fos into stable phosphorylated Fos. ERK/MAPK SLOW branch activates a TF to transcribe the FOS1 gene where FOS1 mRNA is translated into an unstable FOS protein. If the input is not sustained then ERK/MAPK is inactive by the time FOS is transcribed, so the FOS protein simply degrades and we see no output of FOS-mediated transcription.
{kind=link}
Want to create your own Notes for free with GoConqr? Learn more.